51. Protein Digestion: Mechanisms, Enzymes, and Absorption
A perfectly cooked steak or a nutrient-dense lentil stew offers little value to the human body without a microscopic disassembly line. The following slide deck explores the exact mechanisms of this cellular deconstruction. Its core purpose is to break down the highly complex biochemical structures and enzymatic pathways responsible for nutrient absorption. By guiding college and medical students through these pathways, the presentation illuminates how the body masterfully extracts energy and building blocks from dietary intake, revealing the elegant biochemistry that sustains human life.
1. Protein Digestion Overview: How the Body Unlocks Essential Nutrients
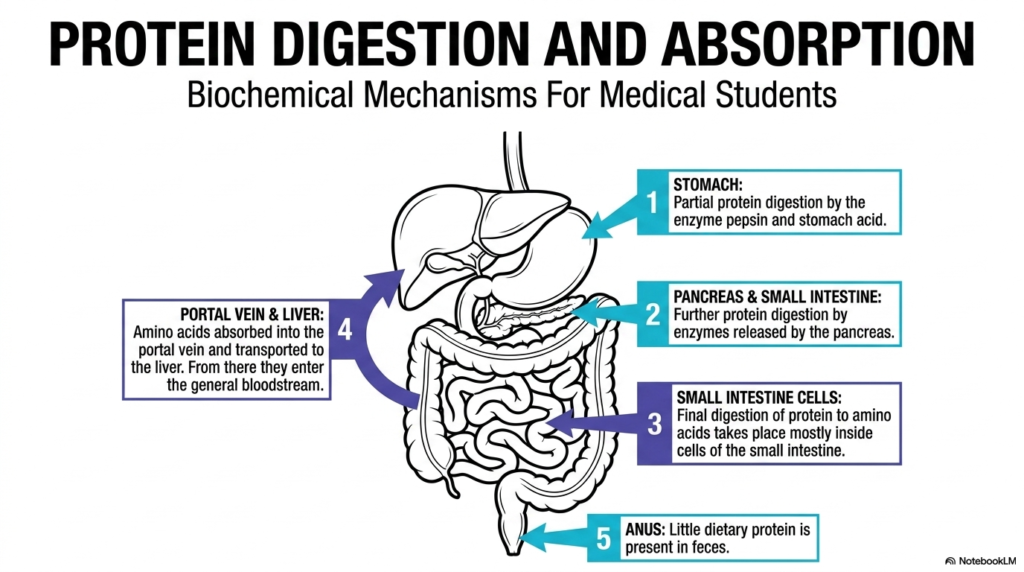
The journey of dietary macromolecules begins with a highly orchestrated physiological event known as Protein Digestion. This initial slide provides a comprehensive topographic map of the entire anatomical pathway that transforms large, complex food items into usable cellular fuel. Students examining this diagram will recognize that the process is never localized to a single organ. Instead, it relies on a sequential, multi-organ collaboration that spans the entire gastrointestinal tract. The stomach, pancreas, small intestine, and liver each play distinct, irreplaceable roles in this metabolic disassembly line. Understanding this macro-level view of anatomy provides the essential framework needed before exploring complex molecular reactions.
Moving chronologically through the diagram, the preliminary phase of Protein Digestion occurs within the highly acidic environment of the stomach. Here, initial unfolding and cleavage take place under extreme physiological conditions. Subsequently, the pathway highlights the pancreas and the small intestine, which collectively introduce a suite of specialized enzymes designed to further dismantle these robust molecular chains. The most critical yet often overlooked final stages occur precisely at the cellular membrane of the small intestine. Specialized intestinal cells, known as enterocytes, carry out the final breakdown into single, absorbable units. The graphic emphasizes the sequential nature of this biological journey.
Once the small intestine completes its enzymatic duties, the newly liberated amino acids must enter the systemic circulation. The final step of Protein Digestion highlighted in this overview involves the transport of these monomers into the portal vein. From this vital vascular highway, the nutrients are directed straight to the liver, the body’s primary metabolic hub, before entering the general bloodstream. Finally, the diagram notes that very little dietary material escapes this highly efficient extraction process, leaving minimal remnants in the feces. This remarkable efficiency underscores the evolutionary perfection of human nutrient absorption.
2. Protein Digestion Fundamentals: The Structural Anatomy of Amino Acids and Peptides
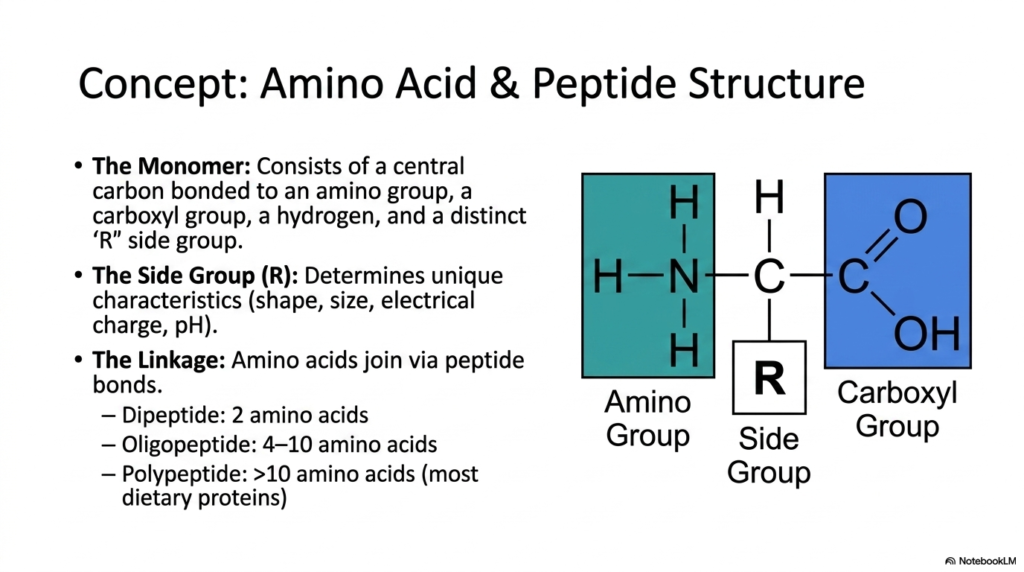
Before comprehending the dynamic enzymatic cleavage involved in Protein Digestion, the medical student must first master the substrate’s structural anatomy. This slide meticulously details the monomeric units of all dietary proteins: individual amino acids. The foundational structure always consists of a central carbon atom covalently bonded to four key components. These include a basic amino group, an acidic carboxyl group, a simple hydrogen atom, and a highly variable ‘R’ side group. The side chain is the defining feature of the molecule, dictating its unique chemical characteristics, physical shape, size, and electrical charge.
The unique properties conferred by these diverse variable side groups are precisely what make the substrate vulnerable to highly specific catalytic enzymes during the sequential stages of Protein Digestion. For instance, some side chains present as bulky, hydrophobic aromatic rings, while others carry strong positive or negative ionic charges that interact dynamically with differing pH environments. When these individual monomers connect to form larger macromolecules, they do so through the formation of robust covalent attachments known as peptide bonds. This specific amide linkage between the carboxyl group of one unit and the amino group of the next is incredibly stable.
Understanding this fundamental linkage is critical, as breaking this exact chemical bond is the ultimate objective of all physiological Protein Digestion. The presentation explicitly categorizes these interconnected biological chains by their structural length for clarity. A dipeptide consists of merely two linked units, while an oligopeptide spans a short chain of four to ten monomers. Polypeptides, which comprise the vast majority of the dietary macronutrients consumed by humans, are large polymeric structures composed of more than 10 covalently linked units. Medical professionals must visualize these dense chains to fully appreciate the monumental biochemical effort required for human nutrition.
3. Gastric Protein Digestion: Acidic Denaturation and Auto-Activation of Enzymes
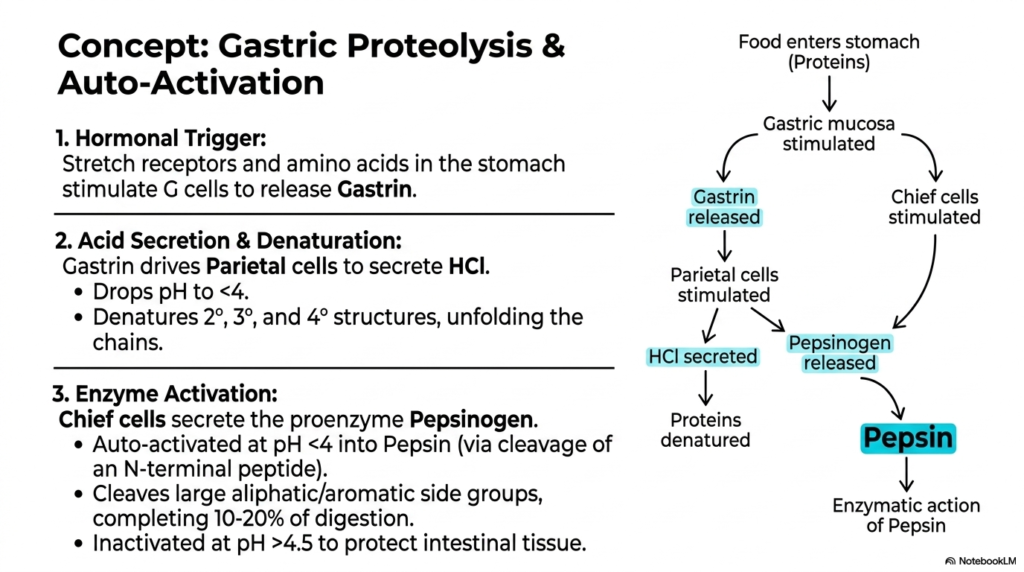
The stomach acts as the initial biochemical battleground for dietary breakdown, launching the first highly active phase of physiological Protein Digestion. This slide introduces the complex hormonal and environmental triggers that orchestrate gastric proteolysis. The sequence begins when physical stretch receptors and the chemical presence of food stimulate specialized G cells within the gastric mucosa. These cells quickly release the hormone Gastrin, which serves as the primary messenger. Gastrin acts upon Parietal cells, driving them to flood the gastric lumen with concentrated hydrochloric acid. This hormonal trigger perfectly synchronizes the structural arrival of food with the body’s powerful chemical secretion mechanisms.
The massive secretion of hydrochloric acid plays an absolutely essential structural role during Protein Digestion by drastically lowering the stomach’s pH to extremely low levels below 4. This highly acidic bath forcefully denatures the complex secondary, tertiary, and quaternary conformations of incoming macromolecules, effectively unfolding the densely packed chains into vulnerable strands. Simultaneously, hormonally stimulated Chief cells secrete an inactive proenzyme, pepsinogen. The true genius of this biological system lies in its brilliant self-regulating auto-activation mechanism. When exposed to harsh acidic conditions, Pepsinogen spontaneously cleaves its N-terminal peptide, thereby transforming into the active protease Pepsin.
Once activated, Pepsin systematically attacks the unwound chains, specifically targeting and cleaving large aliphatic or aromatic side groups. This aggressive enzymatic action manages to complete roughly ten to twenty percent of all necessary Protein Digestion right within the gastric cavity. However, the destructive power of Pepsin must be carefully managed to prevent damage to internal tissues. The slide highlights a fascinating protective mechanism: as the acidic food mass exits the stomach and enters the more alkaline intestinal environment, the rise in pH above 4.5 rapidly inactivates Pepsin. This brilliant evolutionary safeguard completely protects delicate intestinal linings from unwanted autodigestion.
4. Pancreatic Protein Digestion: The Explosive Zymogen Activation Cascade
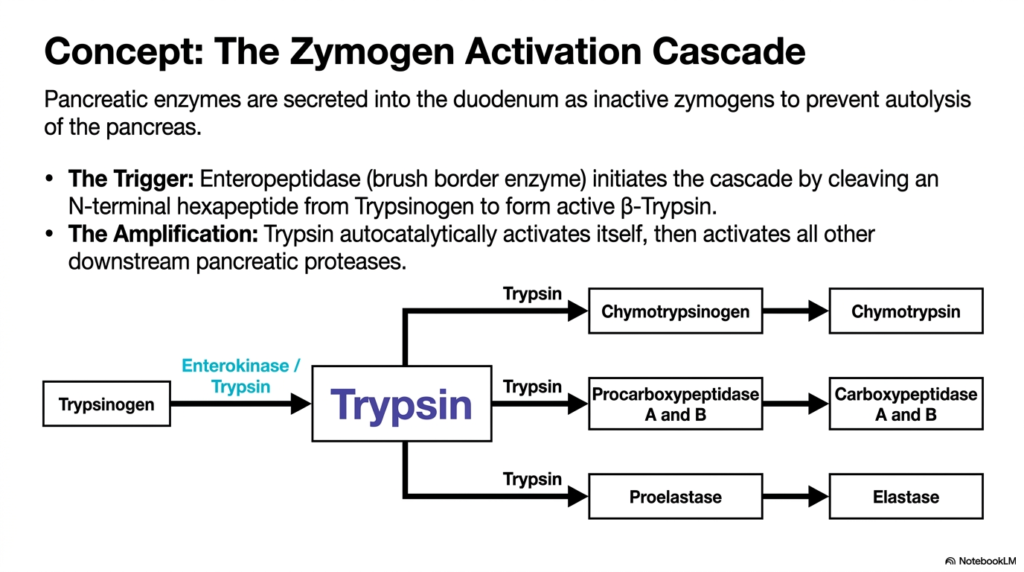
Following gastric processing, the partially degraded mass enters the duodenum, triggering a critical and potentially dangerous phase of Protein Digestion. This slide expertly maps out the zymogen activation cascade, a beautifully designed protective mechanism that prevents the pancreas from digesting itself. The pancreas must produce large quantities of potent proteases, but synthesizing them in their active forms would cause immediate autolysis and severe organ damage. To solve this biological dilemma, the enzymes are secreted as harmless, inactive precursors known as zymogens. They remain entirely dormant during their journey through the pancreatic ducts, waiting for a specific molecular signal.
The spark that ignites this massive enzymatic explosion during Protein Digestion comes from a specialized brush border enzyme called Enteropeptidase, also known as Enterokinase. Located securely on the intestinal wall, Enteropeptidase serves as the highly specific trigger. It carefully cleaves an exact N-terminal hexapeptide from the inactive molecule Trypsinogen, thereby transforming it into the active Beta-Trypsin. This singular event sets off a spectacular biological chain reaction. The newly minted Trypsin exhibits an incredible property: it can autocatalytically activate additional copies of its precursor molecules, creating a massive, localized surge of enzymatic activity within the gut.
With its numbers rapidly multiplying, Trypsin then takes on the role of master conductor for the remainder of luminal Protein Digestion. As illustrated in the detailed flowchart, Trypsin aggressively activates all downstream pancreatic proteases in a swift, coordinated sweep. It directly converts inactive Chymotrypsinogen into the powerful Chymotrypsin, transforms Procarboxypeptidase forms into active Carboxypeptidase A and B, and alters Proelastase into functional Elastase. This cascading amplification ensures that the massive influx of dietary chains is met with an overwhelming, instantaneously generated army of diverse catalytic agents, completely dismantling the dietary structures safely within the intended intestinal lumen.
5. Serine Proteases in Protein Digestion: Unlocking the Catalytic Triad
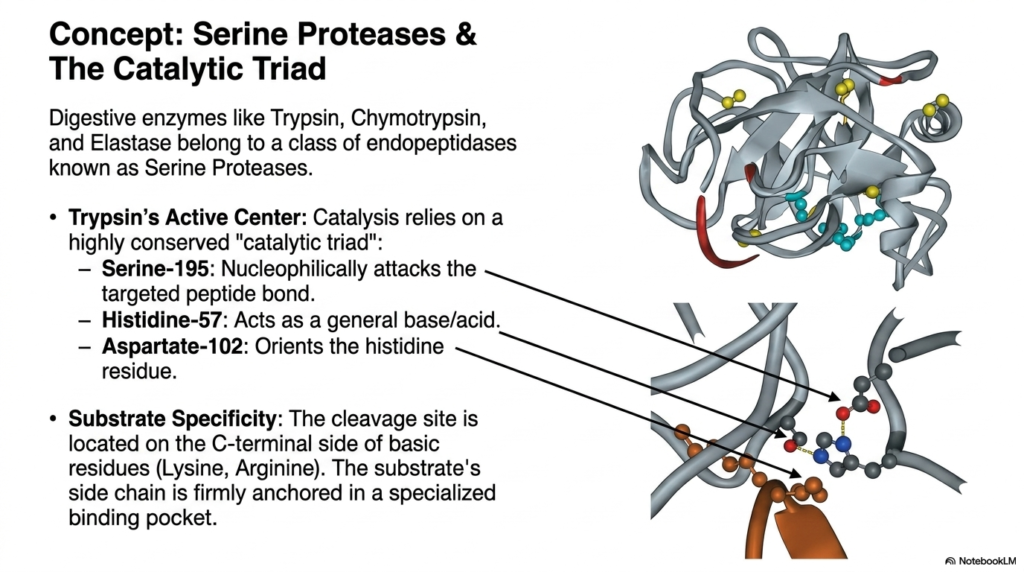
As the activation cascade floods the small intestine with specialized enzymes, the actual molecular machinery of Protein Digestion begins its rapid work. This particular slide delves deep into the structural biochemistry of Serine Proteases, an elite class of endopeptidases that includes Trypsin, Chymotrypsin, and Elastase. The viewer is treated to an intricate visual representation of Trypsin’s active center, showcasing the remarkable precision of molecular evolution. At the heart of this enzyme’s immense catalytic power lies a highly conserved, tightly coordinated structural motif known in biochemistry as the catalytic triad, which is essential for cleaving stubborn amide bonds.
The catalytic triad relies on the perfect three-dimensional alignment of three specific amino acid residues to drive Protein Digestion. Aspartate-102 acts structurally to correctly orient and stabilize Histidine-57 through crucial hydrogen bonding. Histidine-57 then functions as a powerful general base and acid, effectively withdrawing a proton from the incredibly important Serine-195 residue. This biochemical manipulation turns Serine-195 into an exceptionally aggressive nucleophile. The highly reactive oxygen atom on the Serine residue then directly attacks the targeted carbonyl carbon of the substrate’s peptide bond. This coordinated electron transfer represents the exact physical moment at which a protein chain is successfully severed.
Beyond the sheer power of the catalytic triad, successful Protein Digestion requires intense molecular specificity to ensure thorough degradation. The slide expertly illustrates that cleavage sites are not chosen at random. Instead, the enzyme features a highly specialized binding pocket that perfectly anchors the substrate. For Trypsin, this cleavage site is strictly located on the C-terminal side of basic residues, specifically Lysine and Arginine. The physical shape and electrical charge of the binding pocket ensure that only specific peptide sequences fit perfectly against the triad, highlighting the precise, key-and-lock nature of these powerful digestive machines.
6. Endopeptidases vs. Exopeptidases: The Dual Strategy of Protein Digestion
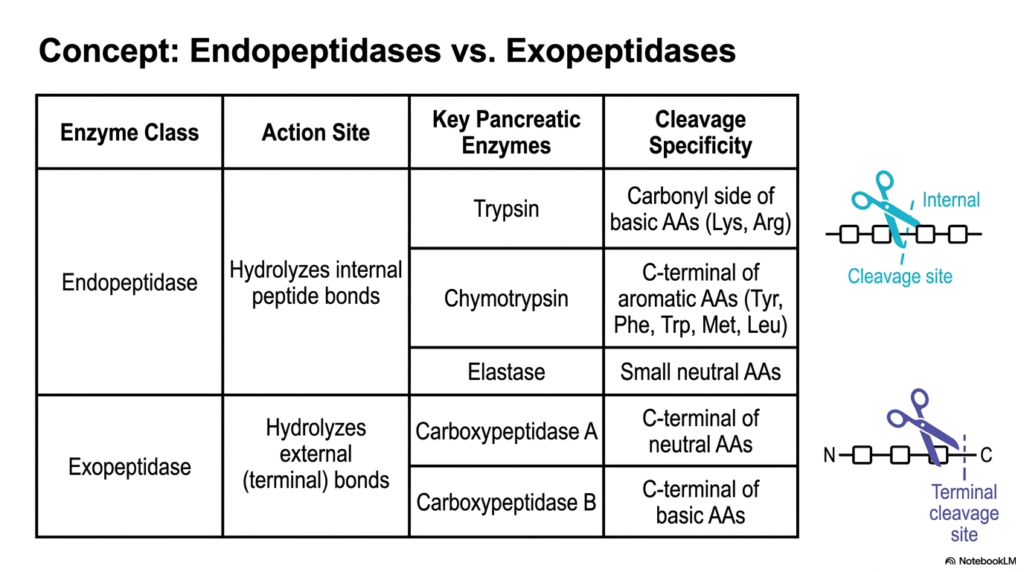
To efficiently dismantle massively complex biological polymers, the gastrointestinal system employs a clever, two-pronged tactical approach during Protein Digestion. This slide provides a foundational comparative analysis between two entirely distinct classes of proteolytic enzymes: Endopeptidases and Exopeptidases. A clear, easy-to-read tabular format categorizes the key pancreatic players by their precise sites of catalytic action. Endopeptidases act like molecular scissors, cutting right into the dense middle of a polypeptide chain, while exopeptidases act like Pac-Men, systematically nibbling away at the very ends of the chain. Both mechanisms are absolutely required for thorough and efficient human nutrient assimilation.
The Endopeptidase class provides the primary bulk-reduction strategy in early luminal Protein Digestion. By targeting internal peptide bonds, enzymes such as Trypsin, Chymotrypsin, and Elastase rapidly fracture massive, unwieldy dietary proteins into significantly smaller, more manageable oligopeptide fragments. As highlighted in the chart, each of these internal cutters possesses unique cleavage specificities. Trypsin favors basic amino acids like Lysine and Arginine, Chymotrypsin hunts for bulky aromatic residues such as Tyrosine and Tryptophan, and Elastase selectively targets small, neutral amino acids. This diverse internal targeting ensures that, regardless of the dietary sequence, the initial long chain is rapidly segmented.
Once the endopeptidases have generated a vast multitude of shorter fragments, the Exopeptidases take command of the final stages of luminal Protein Digestion. These precision enzymes focus exclusively on hydrolyzing the external, terminal bonds of the newly created chains. Carboxypeptidase A systematically cleaves neutral amino acids directly from the exposed C-terminal end, while Carboxypeptidase B specifically targets the available basic amino acids. By relentlessly snipping isolated monomers from the ends of numerous previously fragmented chains, this highly complementary enzymatic system rapidly accelerates the accumulation of free amino acids and highly reduced dipeptides, perfectly preparing the chaotic luminal contents for impending cellular absorption at the intestinal wall.
7. Mucosal and Cytosolic Protein Digestion: The Final Hydrolytic Steps
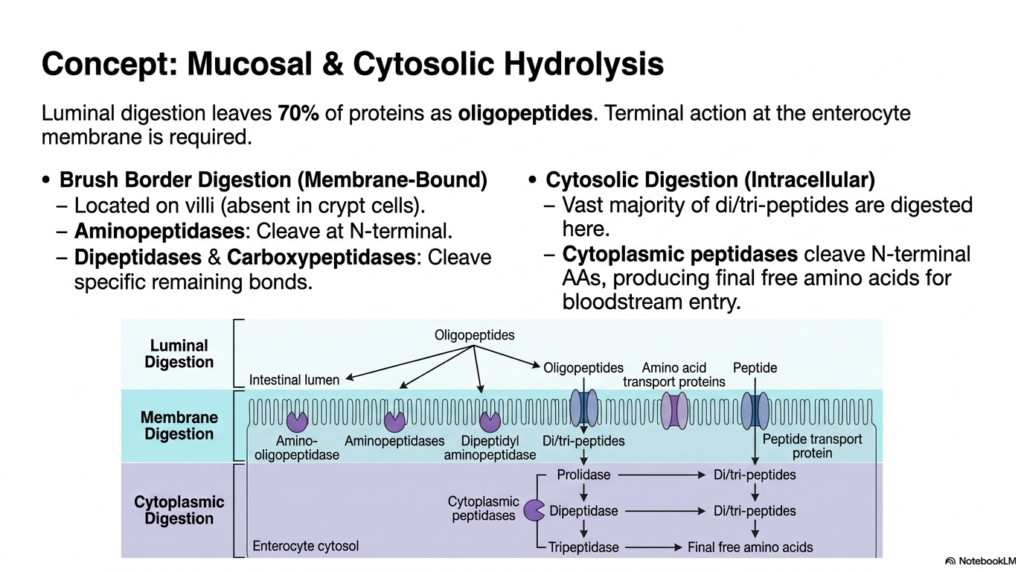
Despite the aggressive action of pancreatic enzymes, bulk luminal breakdown leaves roughly seventy percent of the dietary material as oligopeptides, requiring further Protein Digestion. This fascinating slide shifts the focus away from the chaotic intestinal fluid and directly onto the enterocyte’s highly specialized membrane. To convert these remaining short chains into usable fuel, terminal action is required at the microscopic villi. The brush border of these mucosal cells is densely populated with unique membrane-bound enzymes, which are entirely absent in deeper crypt cells, and is designed to execute the crucial next phase of molecular deconstruction before cellular entry can occur.
The membrane-bound phase of Protein Digestion relies heavily on specialized Aminopeptidases, Dipeptidases, and Carboxypeptidases anchored directly to the intestinal wall. Unlike the earlier pancreatic enzymes, these mucosal tools primarily attack the N-terminal bonds of the approaching oligopeptides. As the molecular fragments brush against the highly folded membrane, they are systematically cleaved into even smaller dipeptides, tripeptides, and free amino acids. The detailed diagram illustrates this literal border crossing, showing how membrane digestion acts as a highly selective biological tollbooth, ensuring that massive, unusable chains are strictly prevented from passing through the enterocyte’s restrictive phospholipid bilayer unprepared.
The final stage of physiological Protein Digestion actually takes place hidden within the cytoplasm of the enterocyte itself. While free amino acids readily pass through, a vast majority of the absorbed material enters the cell as small dipeptides and tripeptides. Within the watery cytosol, highly specialized intracellular peptidases, such as prolidase and tripeptidase, aggressively attack these remaining linkages. This ultimate cytosolic hydrolysis produces the final, unbound free amino acids required for survival. Only after this rigorous, multi-layered intracellular refinement process is complete can the purified nutrients finally be authorized to exit the cell and enter the systemic bloodstream.
8. PEPT1 Transporter in Protein Digestion: High-Speed Peptide Absorption
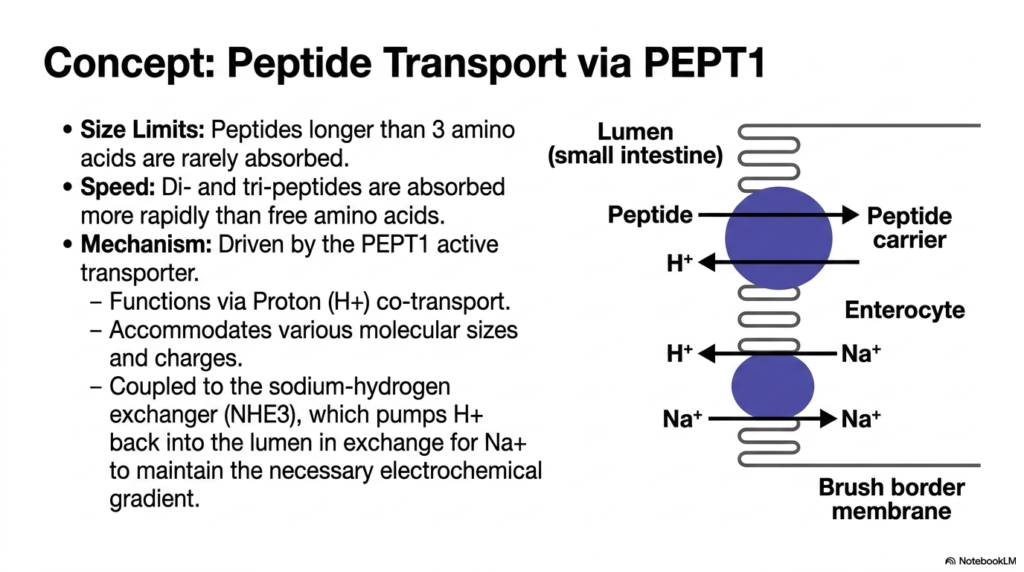
Efficient cellular absorption is just as critical as the chemical cleavage pathways involved in Protein Digestion. This slide highlights a remarkable cellular machinery component, the PEPT1 active transporter, which mediates the rapid uptake of small peptide fragments. The biological constraints of this system are strictly defined by size limits; peptides extending longer than three amino acids are exceptionally rarely absorbed across the intestinal border. However, the system is incredibly efficient for smaller molecules. Remarkably, dipeptides and tripeptides are actually swept into the enterocyte much more rapidly than single, free amino acids, showcasing an evolutionary preference for bulk transport.
The intricate biochemical mechanism driving this high-speed absorption phase of Protein Digestion relies on a brilliantly designed secondary active transport system. The PEPT1 carrier functions specifically via proton (hydrogen ion) co-transport. As illustrated in the detailed schematic, the transporter physically couples the movement of a peptide molecule with the downhill movement of a proton from the lumen into the cell. The immense versatility of the PEPT1 protein structure allows it to accommodate a wide range of molecular sizes and side-chain charges, making it a highly adaptable, universal gateway for nearly all digested di- and tri-peptides.
To maintain the continuous, rapid influx required for optimal Protein Digestion, the cell must cleverly manage its internal environment. The PEPT1 transporter is functionally coupled to a neighboring membrane protein called the sodium-hydrogen exchanger (NHE3). This vital exchanger acts as an internal cellular pump, constantly ejecting the imported protons back out into the intestinal lumen. It accomplishes this task by exchanging the protons for incoming sodium ions, thereby artificially maintaining the steep, necessary electrochemical gradient. This elegant, cyclic interplay of ions ensures that the PEPT1 transporter always has the necessary energetic driving force to continuously absorb dietary peptides.
9. Free Amino Acid Absorption: Secondary Active Transport in Protein Digestion
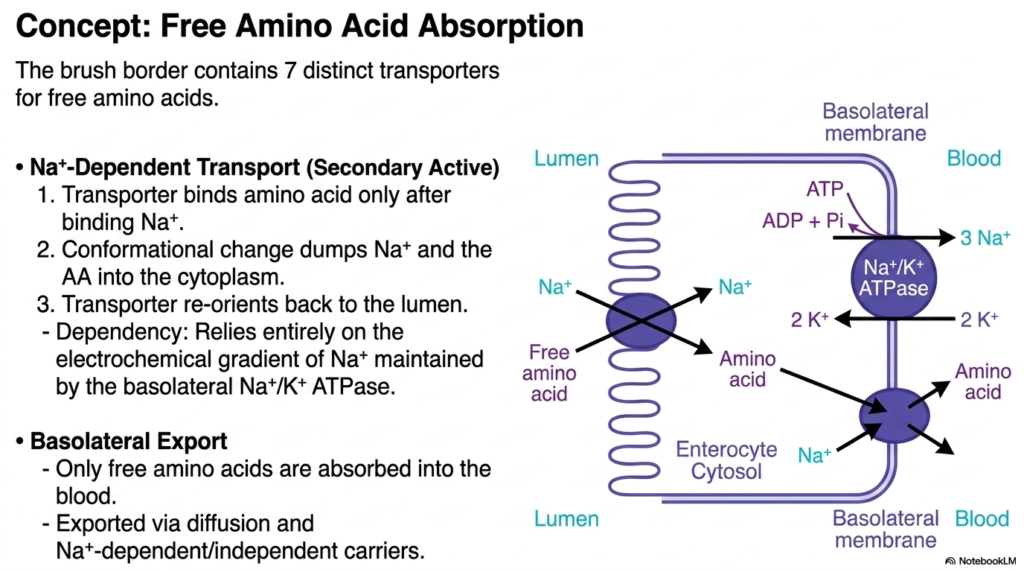
While dipeptides use the PEPT1 gateway, the fully liberated single monomers generated during Protein Digestion require their own dedicated cellular entry pathways. This detailed membrane model outlines the complex absorption pathways for free amino acids. The intestinal brush border is remarkably specialized, containing at least seven distinct, highly specific transporter proteins designed exclusively to ferry these varied individual monomers across the lipid bilayer. The primary mechanism for this vital biological task is sodium-dependent secondary active transport, a highly reliable biological pump that harnesses existing cellular energy gradients to drive nutrients inward against their natural concentration gradients.
The mechanics of this critical stage in Protein Digestion are elegant and highly sequential. First, the empty membrane transporter must securely bind a specific sodium ion from the extracellular fluid. This vital initial binding triggers a crucial conformational change, allowing the target free amino acid to safely attach to the structure. Once fully loaded, the entire protein complex undergoes a massive physical shift, dumping both the sodium ion and the precious amino acid payload deeply into the cellular cytoplasm. The empty transporter immediately reorients itself outward, facing the lumen again, perfectly primed and ready to repeat the continuous absorption cycle.
This entire sodium-dependent phase of Protein Digestion relies completely on the immense power generated by the basolateral sodium-potassium ATPase pump. This unseen cellular engine constantly burns ATP to force sodium out of the cell, maintaining a steep, artificial gradient that makes the apical absorption possible. Once safely inside the enterocyte, the free amino acids face their final biological hurdle: basolateral export. The slide emphasizes that only completely free amino acids are permitted to be absorbed into the systemic blood supply. They seamlessly exit the cell’s rear membrane via simple diffusion and various sodium-independent carriers, finally entering the waiting bloodstream.
10. The Proteasome and Intracellular Protein Digestion: Cellular Quality Control
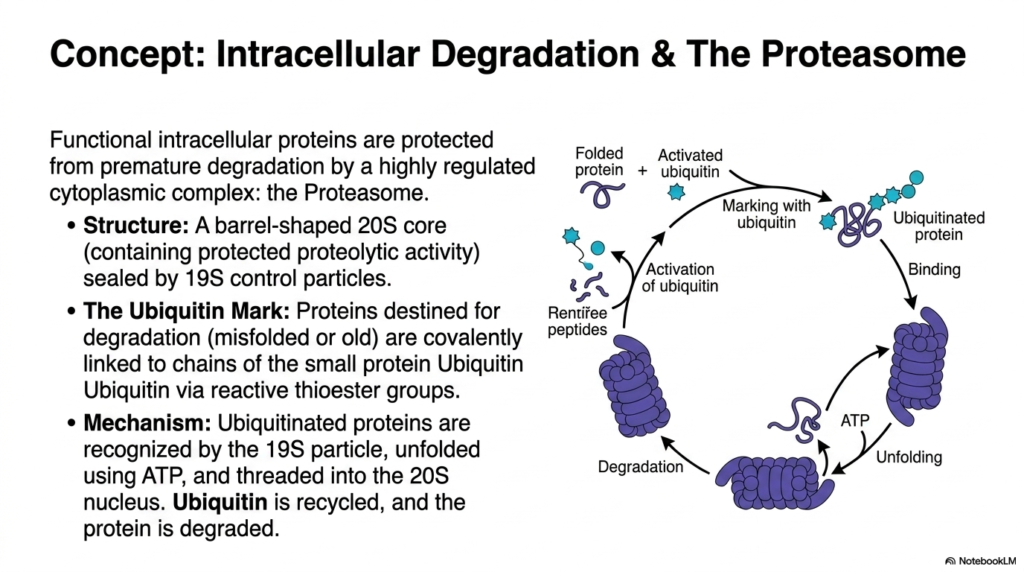
Not all biological breakdown occurs within the digestive tract’s hollow lumen; internal cellular maintenance requires its own highly regulated form of Protein Digestion. This slide pivots entirely from dietary absorption to explore the fascinating realm of intracellular degradation. Within the dense cellular cytoplasm, properly functioning proteins must be aggressively protected from premature destruction. To solve this, the cell isolates its dangerous proteolytic activity safely within a magnificent, barrel-shaped molecular machine known as the Proteasome. This massive 20S core structure is tightly sealed by specialized 19S control particles that strictly regulate molecular entry, acting as highly selective biological bouncers.
To maintain order during this intracellular form of Protein Digestion, the cell utilizes a brilliant chemical tagging system to mark targets for destruction. When internal structural proteins become misfolded, damaged, or simply too old to function, they are chemically condemned. The cell uses reactive thioester groups to covalently link long chains of a small, highly recognizable protein called Ubiquitin directly to the doomed molecule. This visible “Ubiquitin Mark” serves as an undeniable biological death sentence. The complex diagram beautifully illustrates this cyclic tagging pathway, showing how the cell specifically identifies internal structural waste without accidentally destroying its own healthy, functional machinery.
The execution phase of this highly specific Protein Digestion is a marvel of microscopic engineering. The regulatory 19S particle of the Proteasome instantly recognizes the ubiquitous ubiquitin chain and firmly binds the condemned protein. Using massive amounts of cellular energy in the form of ATP, the control particle forcefully unfolds the tangled substrate and physically threads the vulnerable polypeptide chain down into the destructive 20S core nucleus. Inside this secluded biological shredder, the protein is entirely degraded into tiny, harmless peptides. Brilliantly, the precious ubiquitin tags are carefully removed and recycled by the cell, ready to mark the next internal target.
11. Clinical Correlates in Protein Digestion: Genetic Transporter Defects
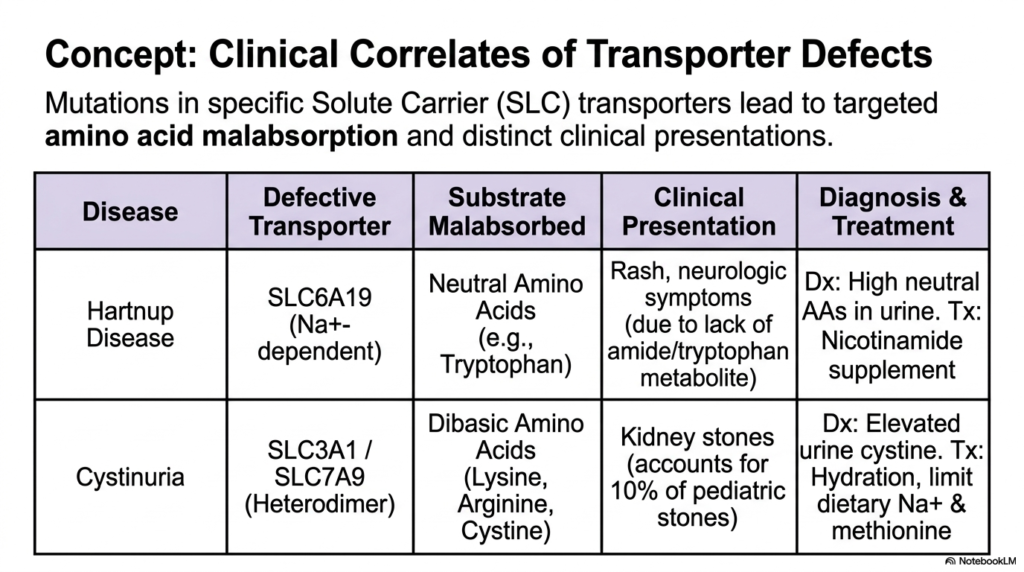
The immense complexity of the molecular pathways governing Protein Digestion means that isolated genetic mutations can lead to profound, highly specific medical pathologies. This slide successfully bridges the gap between pure academic biochemistry and tangible clinical practice by highlighting two devastating genetic diseases directly linked to defects in solute carrier transport. When the intricate membrane proteins responsible for moving individual nutrients fail to function correctly, targeted amino acid malabsorption occurs. The provided table systematically links defective molecular structures directly to their corresponding missing substrates, offering medical students a crystal-clear view of the dangerous systemic consequences of impaired cellular nutrient transport.
The first clinical presentation of failed protein digestion is the fascinating condition known as Hartnup Disease. This specific disorder stems directly from a debilitating genetic mutation in the SLC6A19 sodium-dependent transporter. Because this unique carrier is strictly responsible for moving neutral amino acids, vital nutrients like Tryptophan are completely malabsorbed and subsequently lost in the urine. Clinically, patients present with severe, distinctive skin rashes and dangerous neurologic symptoms. These debilitating systemic issues arise almost entirely because the human body is suddenly starved of crucial amide and tryptophan metabolites, perfectly illustrating the cascading dangers of a single broken transport link.
The second major pathology of impaired Protein Digestion is Cystinuria, a condition fundamentally driven by a devastating mutation in the complex SLC3A1-SLC7A9 heterodimer transporter. This specialized molecular machine typically handles large, dibasic amino acids such as Lysine, Arginine, and Cystine. When this transport pathway fails, the unabsorbed cystine accumulates rapidly and dangerously crystallizes inside the urinary tract, leading to recurrent, highly painful kidney stones. Alarmingly, this specific transport defect accounts for a massive ten percent of all pediatric kidney stone cases. Medical professionals must diagnose these transport failures via elevated urinary amino acids and treat them with intensive hydration protocols.
12. Complete Summary of Protein Digestion: From Stomach to Bloodstream

Consolidating a massive amount of dense biological data, this final slide provides a beautifully synthesized, top-down summary of the entire Protein Digestion pathway. It distills the highly complex, multi-stage biochemical journey into a clear, easily digestible vertical timeline. The visual recap begins at the point where the initial large-scale structural destruction occurs: the highly acidic stomach. Here, intense hydrochloric acid forces structural denaturation, while the aggressive enzyme Pepsin relentlessly cleaves the large, bulky aromatic groups. This foundational step is absolutely necessary to physically prepare the dense macronutrients for the far more delicate and highly precise chemical reactions that await further down the tract.
Following gastric emptying, the summary accurately highlights the intense, localized enzymatic explosion that defines luminal Protein Digestion. As the mass enters the intestine, the mucosal enzyme Enterokinase triggers the massive activation of Trypsin. Trypsin then rapidly unleashes a highly destructive, cascading army of endopeptidases and exopeptidases that viciously fracture the remaining polypeptides. Immediately following this chaotic luminal phase, the organized brush border peptidases take over, meticulously breaking the resulting oligopeptides down into perfectly sized dipeptides, tripeptides, and single amino acids. The clear vertical flowchart expertly reinforces the vital, sequential, and highly ordered nature of these necessary multi-organ enzymatic handoffs.
The ultimate physiological goal of all Protein Digestion is explicitly defined in the final stages of the diagram: absolute cellular absorption and systemic circulation. Enterocytes aggressively use the specialized, high-speed PEPT1 transporters to take up small peptides via proton coupling, while relying on strictly sodium-dependent carriers to safely absorb free monomers. Inside the cell’s watery cytosol, the last remaining cytosolic peptidases forcefully break any lingering bonds, finishing the grueling biological job. The presentation concludes with a stunning metric: over 99% of ingested dietary protein ultimately enters the waiting portal bloodstream as completely purified free amino acids.
Please read our Content Disclaimer Statement.
Check out our social media channels: