70. Plasma Membrane Structure: Lipid Bilayer and Proteins Explained
Every living cell constantly fights a microscopic battle for survival against a chaotic, unpredictable environment. The ultimate defender in this continuous biological struggle is the cellular membrane. This carefully curated slide deck breaks down the incredibly complex biochemical architecture of the cellular boundary into easily digestible, highly visual concepts. By exploring these illustrations, biochemistry students will uncover exactly how specific lipids, massive embedded proteins, and complex carbohydrates seamlessly combine to form a dynamic, selectively permeable shield. Understanding these vital structural mechanisms reveals the foundational molecular rules governing all complex cellular life.
1. Plasma Membrane Structure: Biochemical Architecture
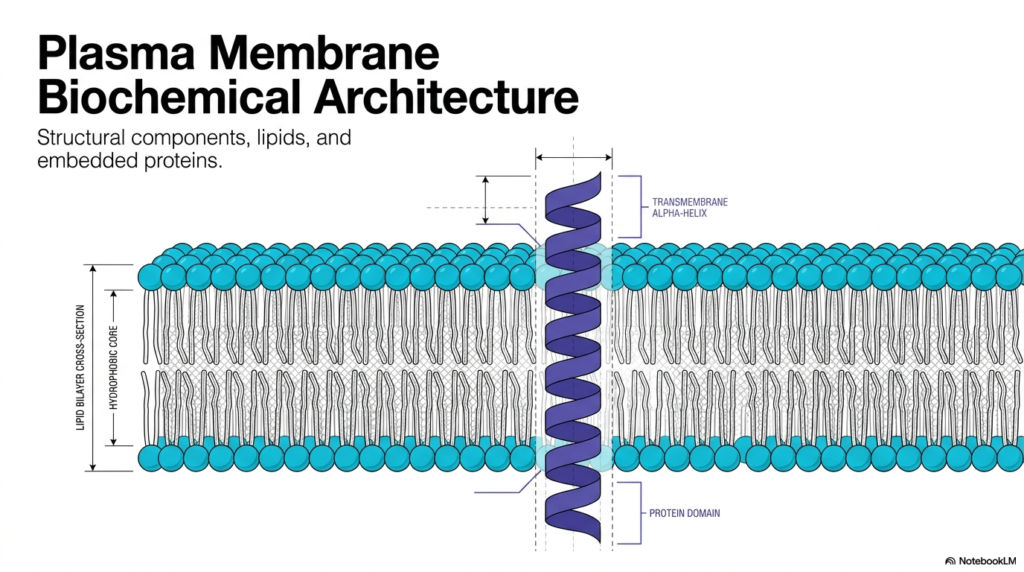
The study of cellular boundaries begins with the foundational biochemical architecture of the cell. Understanding the Plasma Membrane Structure is essential for comprehending how biological cells interact with their surrounding environment. This primary barrier effectively separates internal cellular machinery from the unpredictable external world. The provided visual schematic illustrates a highly organized lipid bilayer cross-section. It highlights a prominent hydrophobic core that inherently restricts the free passage of water-soluble molecules. Within this complex environment, various structural components, distinct lipid species, and embedded proteins function collectively to uphold cellular integrity. This molecular barrier represents a highly selective gateway rather than a static biological wall.
A rigorous examination of the Plasma Membrane Structure reveals the strategic placement of transmembrane alpha-helices. These specialized protein domains completely span the width of the lipid bilayer. The hydrophobic core of the biological membrane interacts favorably with the nonpolar amino acid residues forming the alpha-helix. This thermodynamic interaction firmly anchors the protein within the lipid matrix while enabling it to perform crucial transport or signaling functions. The biochemical equilibrium between the lipid environment and these embedded protein domains remains a fundamental principle of cellular biochemistry. Embedded proteins actively facilitate communication, structural support, and material transport across the cellular boundary.
Mastering the physical intricacies of the Plasma Membrane Structure establishes a solid knowledge base for advanced pharmaceutical and physiological studies. Numerous medical therapies specifically target these embedded transmembrane proteins to modulate cellular behavior and treat diseases. The lipid bilayer itself measures merely a few nanometers in thickness, yet it provides extraordinary mechanical stability to the organism. Analyzing the transmembrane alpha-helix securely anchored within the hydrophobic core demonstrates the remarkable elegance of cellular design. The structural components depicted in this first diagram form the universal biological blueprint for all living cells, initiating a comprehensive exploration of molecular cell biology.
2. Plasma Membrane Structure: The Fluid Mosaic Architecture
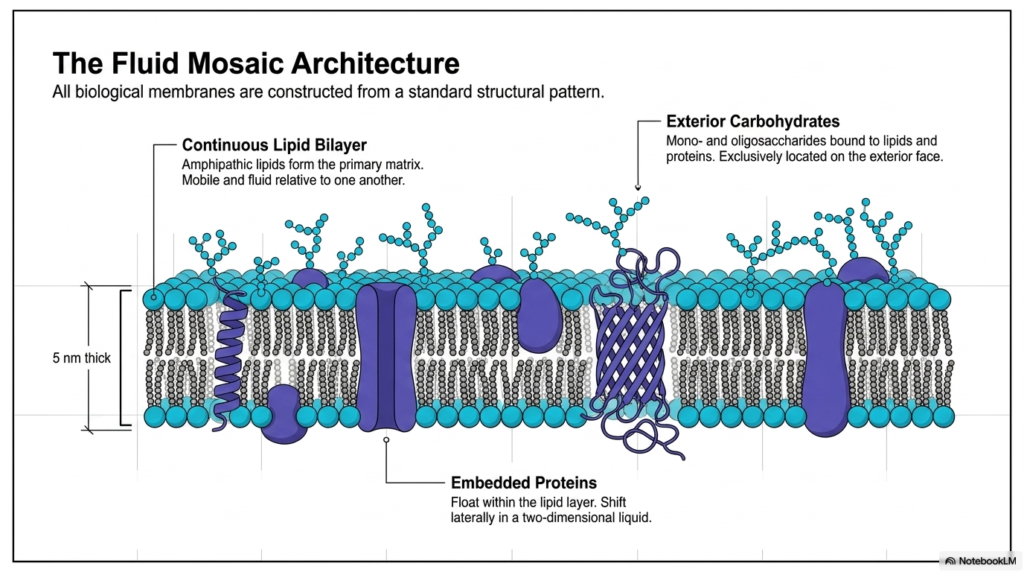
The widely accepted model for biological boundaries relies heavily on the fluid mosaic architecture. Examining the Plasma Membrane Structure through this lens reveals a continuous lipid bilayer that forms the primary matrix of the cell. Measuring exactly five nanometers thick, this delicate layer consists of amphipathic lipids that remain highly mobile and fluid relative to one another. The model accurately portrays biological membranes as a dynamic, two-dimensional liquid rather than a rigid structural solid. Embedded proteins float freely within this lipid sea, shifting laterally to perform varied biochemical tasks. This specific architectural pattern remains standard across countless living organisms.
Another vital aspect of the Plasma Membrane Structure involves the exterior carbohydrates strategically positioned on the outer leaflet. Mono- and oligosaccharides covalently bind to underlying lipids and proteins, functioning exclusively on the exterior face of the cell. These specialized sugar chains assist in cellular recognition, adhesion, and protection against mechanical stress. The combination of the continuous lipid bilayer and these projecting carbohydrate structures creates a highly complex cellular surface. Integral and peripheral embedded proteins continuously navigate this dense matrix. Their lateral movement within the two-dimensional liquid enables dynamic biological responses to changing environmental conditions.
The brilliance of the Plasma Membrane Structure lies in its ability to combine diverse biochemical components into one cohesive, functional unit. The continuous lipid layer provides the essential hydrophobic barrier, while the embedded proteins operate as specialized functional machines. Exterior carbohydrates add a crucial layer of chemical identity to the cell surface. Together, these molecules form a dynamic mosaic that shifts and adapts instantly. The five-nanometer thickness belies the immense complexity and resilience of the biological boundary. This fluid mosaic configuration ultimately allows cells to constantly maintain chemical homeostasis while efficiently communicating with neighboring biological entities.
3. Plasma Membrane Structure: Amphipathic Assembly Mechanics
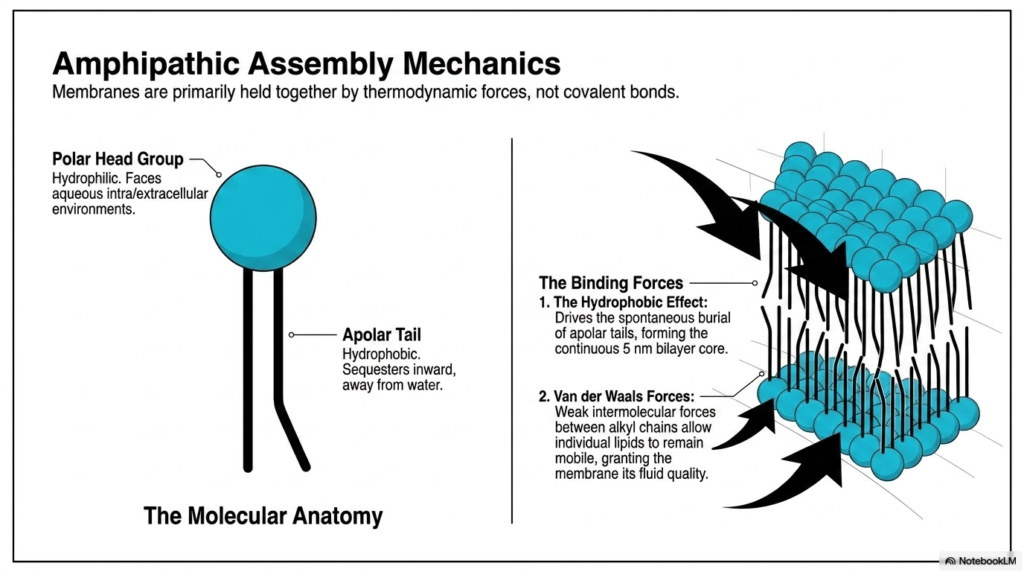
The spontaneous formation of biological boundaries relies on precise amphipathic assembly mechanics. The core of the Plasma Membrane Structure forms primarily through distinct thermodynamic forces rather than covalent bonds. Each membrane lipid exhibits a unique molecular anatomy, featuring a hydrophilic polar head group and a hydrophobic apolar tail. The polar head actively faces the aqueous intracellular and extracellular environments, eagerly interacting with water molecules. Conversely, the apolar tail sequesters itself inward to escape the aqueous surroundings. This fundamental structural dichotomy perfectly drives the spontaneous assembly of the continuous five-nanometer bilayer core. Molecules with both hydrophilic and hydrophobic regions represent the ultimate building blocks of cellular biology.
Understanding the powerful driving forces behind the Plasma Membrane Structure requires analyzing both the hydrophobic effect and Van der Waals forces. The hydrophobic effect is the primary molecular driver, driving the spontaneous burial of apolar tails to minimize highly unfavorable interactions with surrounding water molecules. This powerful thermodynamic phenomenon pushes the individual lipids to aggregate aggressively, creating a highly stable, continuous matrix. Once the core is successfully assembled, the Plasma Membrane Structure relies heavily on weak intermolecular Van der Waals forces between adjacent alkyl chains. These delicate interactions keep the lipids associated while remaining weak enough to confer the membrane’s signature fluid quality.
The resulting molecular anatomy guarantees that the boundary remains both highly stable and biologically flexible. Without these powerful thermodynamic forces, the lipid bilayer would instantly dissolve into the surrounding aqueous environment. The careful biochemical balance between the hydrophilic polar head group, which seeks water, and the hydrophobic apolar tail entirely dictates the architecture. The continuous five-nanometer-thick bilayer core provides an impenetrable shield against charged ions and large polar molecules. By relying solely on spontaneous amphipathic assembly, the entire Plasma Membrane Structure efficiently conserves valuable chemical energy while rapidly constructing its most vital biological barrier. This robust self-assembling nature intelligently ensures incredibly rapid membrane repair.
4. Plasma Membrane Structure: Thermodynamics of Fluidity
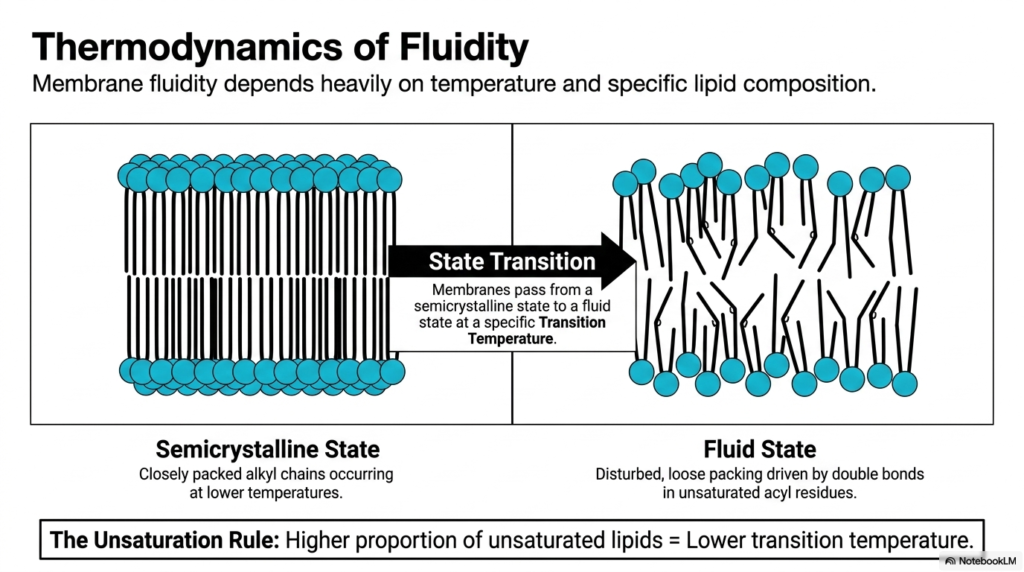
The physical state of cellular boundaries depends intricately on the thermodynamics of fluidity. The Plasma Membrane Structure actively transitions between different physical states based on environmental temperature and specific lipid composition. At lower temperatures, the membrane adopts a semicrystalline state characterized by closely packed alkyl chains. This highly ordered configuration restricts molecular movement and rigidifies the cellular boundary. However, biological membranes must pass from this semicrystalline state to a functional fluid state at a specific transition temperature. Maintaining optimal fluidity is absolutely vital for the proper functioning of embedded transport proteins and cellular signaling cascades.
The specific lipid composition deeply influences how the Plasma Membrane Structure responds to thermal changes. The Unsaturation Rule states that a higher proportion of unsaturated lipids directly results in a lower transition temperature. Unsaturated acyl residues contain distinct double bonds that create physical kinks in the hydrocarbon chains. These prominent kinks prevent the lipids from packing too tightly together, creating a disturbed, loose packing arrangement. This crucial biochemical variation guarantees that the Plasma Membrane Structure remains in a highly mobile fluid state even when environmental temperatures drop significantly. The physical disturbance caused by these double bonds ensures that biological membranes avoid freezing solid under standard physiological conditions.
Organisms constantly modify their specific lipid composition to survive wildly fluctuating thermal environments. By increasing the incorporation of unsaturated acyl residues, cells actively lower their transition temperature and prevent dangerous membrane solidification. The fluid state allows individual lipid molecules and embedded proteins to diffuse rapidly across the biological surface. Without this essential thermodynamic flexibility, cellular transport mechanisms would immediately halt, leading to rapid cell death. The delicate balance between the closely packed semicrystalline state and the disturbed fluid state represents a masterful biochemical adaptation. This dynamic state transition ensures the Plasma Membrane Structure maintains continuous cellular survival across diverse ecological environments.
5. Plasma Membrane Structure: Cholesterol, the Membrane Buffer
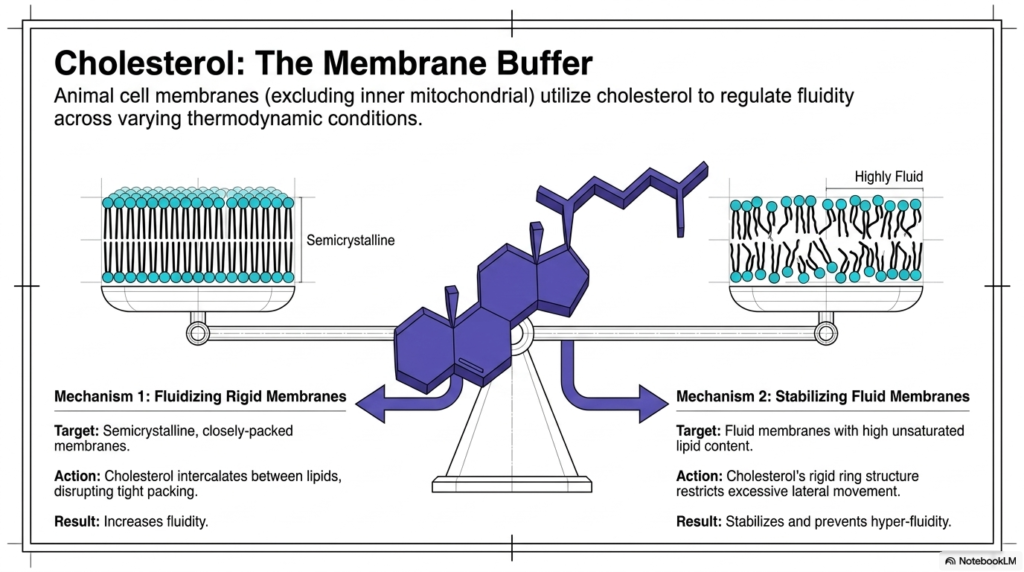
Animal cell membranes prominently utilize cholesterol to carefully regulate fluidity across varying thermodynamic conditions. Within the Plasma Membrane Structure, cholesterol acts as an essential bidirectional buffer of fluidity. Interestingly, the inner mitochondrial membrane lacks this molecule, but it remains abundant throughout the primary animal cell boundary. The rigid ring structure of cholesterol allows it to actively modulate the physical properties of the lipid bilayer. It effectively manages the delicate balance between the semicrystalline and highly fluid states. Without this vital lipid buffer, animal cells would struggle to maintain structural integrity during extreme temperature shifts.
Cholesterol executes its buffering role within the Plasma Membrane Structure through two highly distinct biochemical mechanisms. The first mechanism involves fluidizing rigid, closely packed semicrystalline membranes. Cholesterol physically intercalates between the tightly bound lipid molecules, effectively disrupting their tight packing. This specific intercalating action forcefully prevents the alkyl chains from crystallizing, thereby increasing overall membrane fluidity. The Plasma Membrane Structure actively relies on this disruption to remain pliable and functional during sudden temperature decreases. This fluidizing action guarantees that the cellular surface does not become rigidly locked or biologically inert.
The second functional mechanism involves stabilizing highly fluid membranes that contain extensive unsaturated lipid content. In this scenario, the rigid ring structure of cholesterol chemically restricts excessive lateral movement among the rapidly shifting lipids. This stabilizing action successfully prevents the boundary from achieving dangerous hyper-fluidity, which would ultimately compromise essential cellular containment. By simultaneously preventing solid crystallization and restricting chaotic hyper-fluidity, cholesterol perfectly fine-tunes the physical dynamics of the Plasma Membrane Structure. This remarkable biochemical molecule ensures that the biological barrier maintains optimal transport function and mechanical strength despite fluctuating external thermodynamic pressures. The bidirectional nature of this lipid buffer represents a pinnacle of evolutionary biochemistry.
6. Plasma Membrane Structure: Lateral vs. Transverse Movement
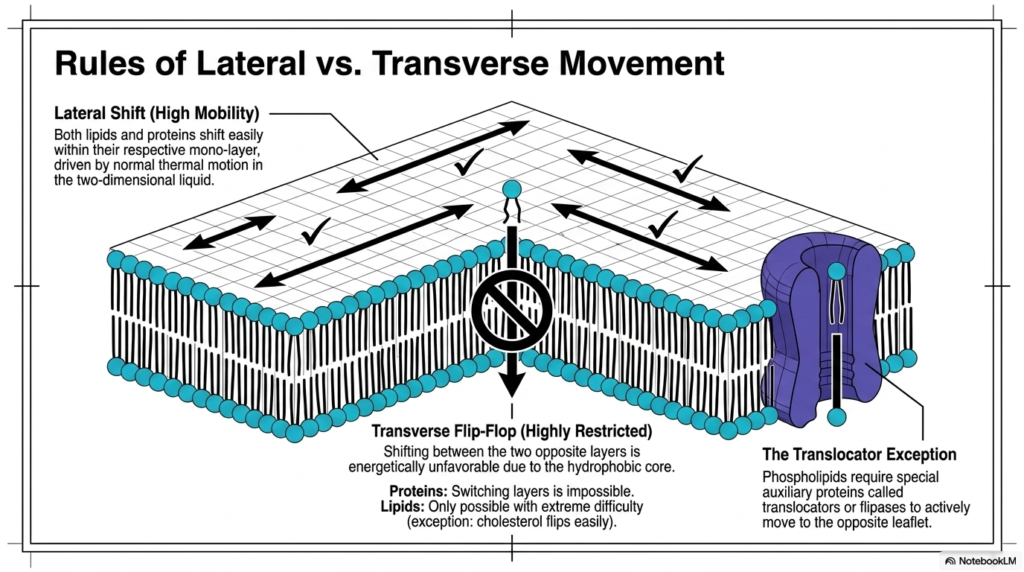
The dynamic nature of the cellular boundary follows strictly established physical rules governing molecular mobility. Within the Plasma Membrane Structure, individual molecules experience vastly different degrees of freedom depending solely on their precise direction of travel. Lateral shift represents a continuous state of high mobility, allowing both lipids and proteins to move freely within their respective monolayers. This rapid horizontal movement is constantly driven by normal ambient thermal motion actively occurring within the two-dimensional liquid matrix. The unhindered lateral shifting continuously enables diverse proteins to locate one another, physically interact, and successfully execute complex biochemical signaling cascades.
Conversely, transverse flip-flop motion within the Plasma Membrane Structure is severely constrained by strong thermodynamic constraints. Shifting between the two opposite layers proves energetically unfavorable because the hydrophilic polar head must forcibly drag itself through the thick hydrophobic core. For massive embedded proteins, switching layers is physically impossible and permanently forbidden. For individual lipid molecules, this transverse flip-flop remains possible only with extreme difficulty, though small molecules like cholesterol can flip rather easily. The Plasma Membrane Structure inherently strongly resists this spontaneous flipping to carefully maintain the established biochemical asymmetry of the two leaflets.
To intelligently overcome this severe physical restriction, the biological system consistently employs a remarkable translocator exception. Highly specialized auxiliary proteins, specifically called translocators or flipases, actively move selected phospholipids to the opposite leaflet. These vital transmembrane enzymes frequently utilize cellular energy to safely transport the sensitive polar head groups across the hostile hydrophobic core. This targeted, enzyme-driven movement establishes and rigorously maintains the highly distinct lipid compositions critically required on each unique side of the cellular boundary. The translocators actively bypass the physical constraints of the Plasma Membrane Structure to securely transport lipids and guarantee proper spatial organization.
7. Plasma Membrane Structure: Phospholipids as the Primary Matrix
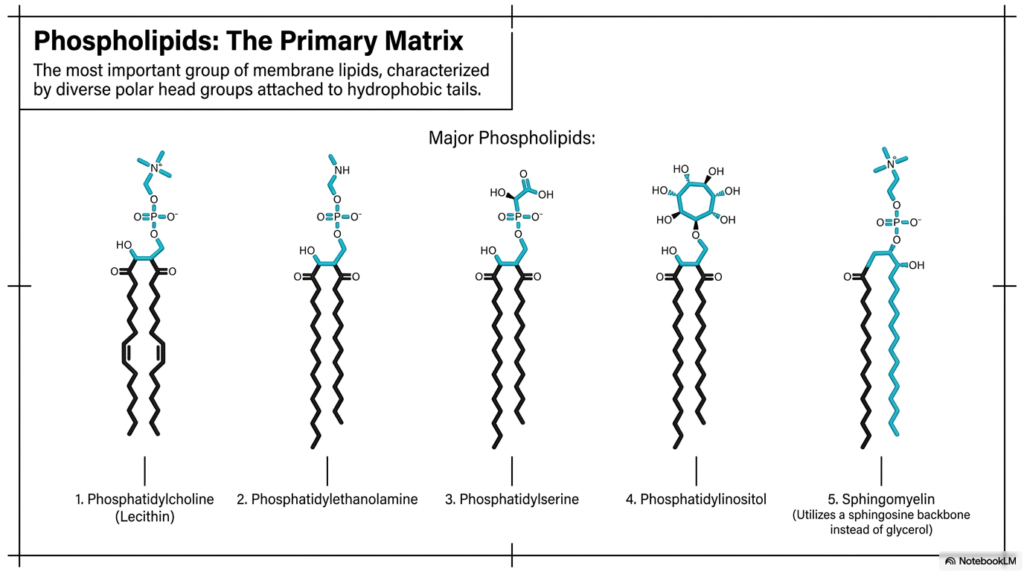
The ultimate structural foundation of the cellular boundary relies entirely on specific classes of highly complex lipids. The single most important group of lipids directly forming the Plasma Membrane Structure is the phospholipids. These essential biochemical molecules universally function as the primary matrix of the cell. They are structurally characterized by diverse, highly reactive polar head groups covalently attached to long, inert hydrophobic tails. These unique lipid molecules spontaneously organize into the continuous five-nanometer bilayer that distinctly defines all known biological boundaries. Their inherent amphipathic nature perfectly dictates the fundamental physical properties of the entire cellular surface.
Five major phospholipids prominently dominate the complex biochemical composition of the Plasma Membrane Structure. Phosphatidylcholine, commonly known in biochemistry as Lecithin, is typically the most abundant phospholipid in the exterior leaflet. Phosphatidylethanolamine and Phosphatidylserine typically concentrate heavily along the inner cytoplasmic leaflet, critically contributing essential negative charges to the internal cellular environment. Phosphatidylinositol simultaneously plays a uniquely critical role in directing complex intracellular signaling cascades despite being present in remarkably smaller quantities. The highly diverse, asymmetrical distribution of these specific lipid molecules throughout the Plasma Membrane Structure actively creates a highly specialized biological barrier.
The fifth and final major matrix component is Sphingomyelin, which noticeably differs structurally from the other primary phospholipids. Instead of a standard glycerol backbone, Sphingomyelin uses a distinct sphingosine backbone to securely anchor its hydrophobic tails. These beautifully subtle biochemical variations among the five major phospholipids explicitly grant the Plasma Membrane Structure entirely unique mechanical and chemical characteristics. The careful biological synthesis and exact spatial distribution of these distinct molecular species allow the living cell to precisely tailor its barrier properties. A comprehensive understanding of these primary matrix components remains absolutely fundamental for mastering advanced cellular biochemistry.
8. Plasma Membrane Structure: The Glycocalyx Cellular Coating
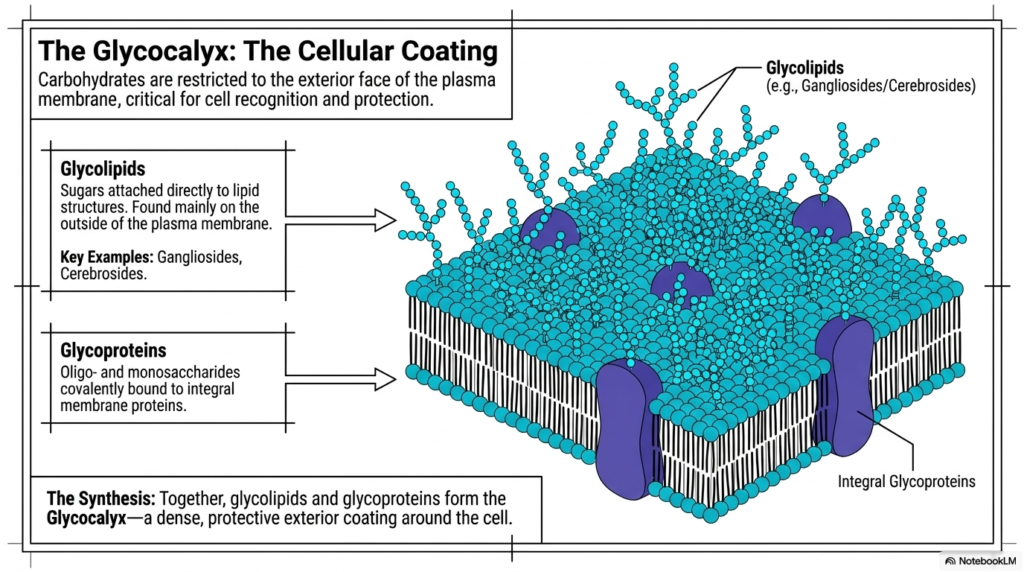
The exterior face of the cellular boundary features a highly specialized, remarkably dense protective chemical coating. This vital exterior modification of the Plasma Membrane Structure is scientifically known throughout biology as the Glycocalyx. Crucially, functional carbohydrates are strictly restricted to the exterior face of the biological barrier, remaining completely absent from the internal cytoplasmic leaflet. This dense, highly complex sugar coating proves absolutely critical for advanced cell recognition, immune system navigation, and robust physical protection against mechanical damage. The intricate, branching network of projecting carbohydrates completely transforms the otherwise smooth lipid bilayer into an interactive biological landscape.
The formation of the Glycocalyx depends heavily on two major molecular classes embedded within the Plasma Membrane Structure. The first class, glycolipids, consists of complex sugars attached directly to structural lipid molecules. These unique glycolipids, which notably include vital examples like Gangliosides and Cerebrosides, are found almost exclusively on the outside of the cellular boundary. The second major structural class involves glycoproteins. These consist of specialized oligo- and monosaccharides covalently bound directly to large integral membrane proteins. The Plasma Membrane Structure utilizes these massive glycosylated proteins to physically interact with the surrounding extracellular matrix and neighboring cells.
Together, these exceptionally diverse glycolipids and massive integral glycoproteins effectively synthesize the continuous protective Glycocalyx. This dense, noticeably sticky exterior coating successfully traps surrounding water molecules, actively creating a highly slippery physical barrier around the entire living cell. This specialized biochemical hydration layer prevents mobile cells from inappropriately adhering to one another while they rapidly flow through narrow circulatory vessels. Furthermore, the unique superficial carbohydrate sequencing serves as a highly specific biochemical fingerprint, enabling the immune system to reliably identify friendly native cells. The Glycocalyx perfectly highlights how the Plasma Membrane Structure incorporates complex carbohydrate chemistry to ensure absolute cellular survival.
9. Plasma Membrane Structure: Protein Integration Mechanisms
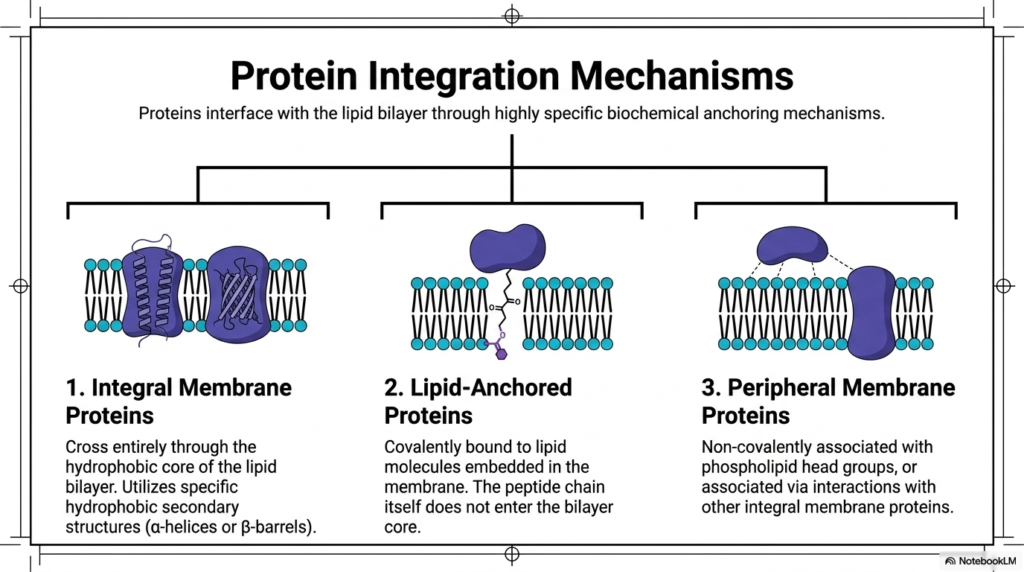
The true functional capacity of the cellular boundary relies entirely on carefully embedded, highly specialized protein machines. Crucial proteins securely interface with the Plasma Membrane Structure through highly specific and remarkably complex biochemical anchoring mechanisms. Without these diverse and necessary integration strategies, vital proteins would simply wash away uselessly into the surrounding aqueous environment. The biological membrane actively employs three distinct structural categories to associate with essential functional proteins. Comprehensively understanding these distinct physical integration mechanisms is essential for mastering how living cells transport materials, respond to external chemical signals, and mechanically bind to neighboring tissues.
The single most prominent integration mechanism found within the Plasma Membrane Structure involves massive Integral Membrane Proteins. These complex molecular structures completely cross the hostile, water-repelling hydrophobic core of the lipid bilayer. To survive this highly thermodynamically unfavorable environment, they use specific hydrophobic secondary structures, primarily alpha-helices and antiparallel beta-barrels. Alternatively, Lipid-Anchored Proteins associate with the Plasma Membrane Structure through distinct biochemical mechanisms. These unique functional proteins are firmly covalently bound to specific lipid molecules already deeply embedded in the continuous membrane matrix. Crucially, the functional peptide chain itself never actively enters the dangerous hydrophobic bilayer core.
The third and final major integration category explicitly consists of highly dynamic Peripheral Membrane Proteins. These specialized molecules completely refuse to penetrate the hydrophobic core and never utilize direct covalent lipid attachments. Instead, they remain strictly non-covalently associated with the negatively charged phospholipid head groups safely located along the aqueous boundary. Furthermore, they frequently associate via strong physical interactions directly with other permanently established integral membrane proteins. These peripheral proteins can rapidly and transiently attach and detach as needed, thereby vigorously driving dynamic intracellular signaling cascades. Together, these three highly sophisticated integration mechanisms perfectly populate the Plasma Membrane Structure.
10. Plasma Membrane Structure: Integral alpha-Helical Anchors
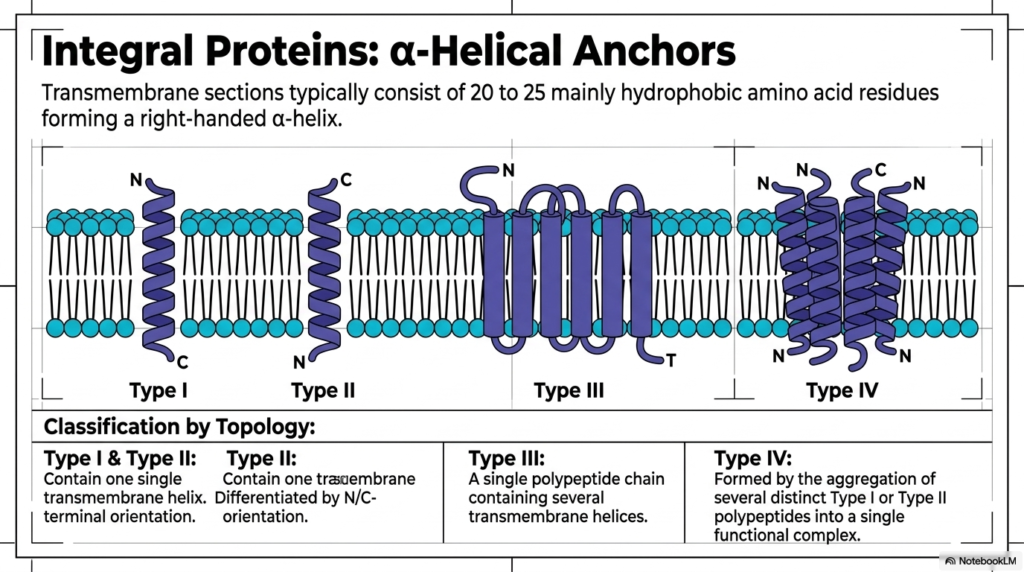
The single most common structural mechanism by which massive proteins physically penetrate the cellular boundary involves entirely distinct helical structures. Deep within the Plasma Membrane Structure, critical integral proteins primarily utilize highly stable alpha-helical anchors to safely span the dangerous hydrophobic core. These highly specialized transmembrane sections typically consist of precisely twenty to twenty-five mainly hydrophobic, nonpolar amino acid residues. By naturally adopting a tight, right-handed alpha-helix physical conformation, the embedded protein successfully shields its delicate internal polar peptide bonds from the hostile, water-hating lipid environment. This elegant biochemical solution provides immense structural stability, allowing massive receptor complexes to remain securely locked within the biological boundary.
The strict scientific classification of these essential alpha-helical anchors within the Plasma Membrane Structure depends heavily on their complex molecular topology. Both Type I and Type II integral proteins contain a single continuous transmembrane helix and function as simple single-pass structural anchors. However, these two types are highly differentiated by their specific N- and C-terminal orientations directly across the dense lipid bilayer. The Plasma Membrane Structure actively and rigorously maintains this strict orientational asymmetry, ensuring that exterior receptor domains and interior signaling domains remain perfectly positioned. A reversed biochemical orientation would completely destroy the specific functional capacity of the embedded transmembrane protein.
Significantly more complex biological integration requirements absolutely rely on highly advanced multi-pass protein architectures. Type III integral membrane proteins consist of a single, highly elongated polypeptide chain specifically containing several distinct transmembrane helices woven seamlessly together. These multiple integrated helices frequently bundle closely together to effectively form sophisticated internal transport channels or massive sensory receptors. Alternatively, massive Type IV integral structures are physically formed by the careful physical aggregation of several distinct Type I or Type II individual polypeptides. These multiple independent peptide chains assemble highly cooperatively to actively form a single, incredibly massive functional complex. This remarkably diverse array of specific alpha-helical topologies allows the Plasma Membrane Structure to actively construct customized molecular machinery.
11. Plasma Membrane Structure: The beta-Barrel Architecture
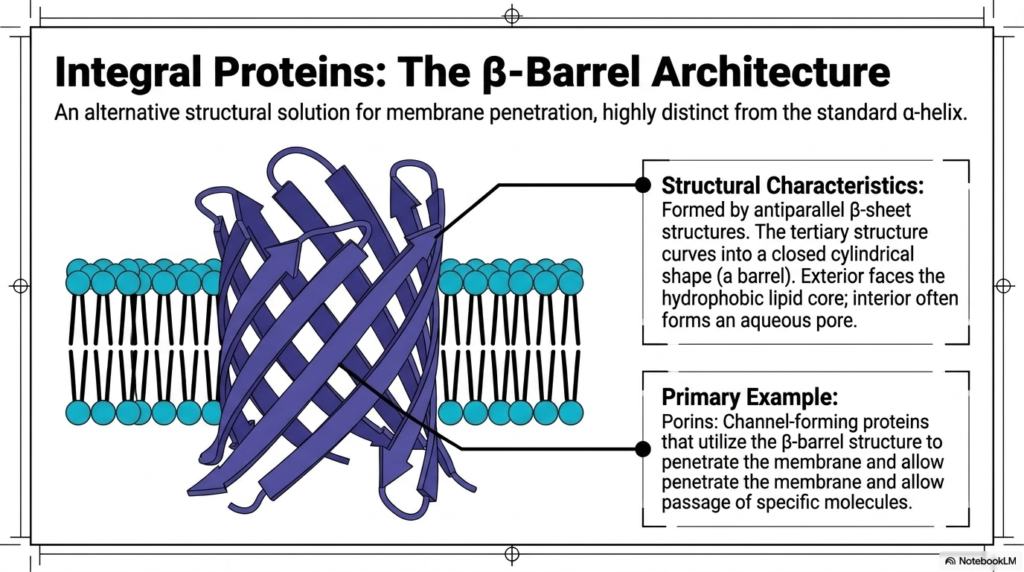
While standard helical structures heavily dominate, biological systems possess brilliant alternative methods for crossing the cellular boundary. The Plasma Membrane Structure occasionally utilizes a radically different approach known as the beta-barrel architecture. This structural solution for direct membrane penetration remains highly distinct from the ubiquitous right-handed alpha-helix. Instead of a tightly coiled spiral, this massive architecture is meticulously formed by multiple elongated antiparallel beta-sheet structures. These extended biological sheets physically curve and tightly fold into a completely closed, rigid cylindrical shape, creating a hollow molecular barrel. This fascinating structural configuration is primarily found embedded within the outer boundaries of bacteria, mitochondria, and chloroplasts.
The true biochemical brilliance of the rigid beta-barrel explicitly found within the Plasma Membrane Structure heavily relies on its highly specific spatial orientation of selected amino acids. The strictly alternating molecular sequence of the folded antiparallel beta-sheets perfectly dictates that the barrel’s exterior surface entirely faces the hostile hydrophobic lipid core. Consequently, these specific exterior amino acids are strictly nonpolar and extremely hydrophobic. Conversely, the hollow interior of the folded beta-barrel predominantly features highly reactive hydrophilic amino acid residues. This specific Plasma Membrane Structure arrangement intelligently allows the hollow interior to successfully form a continuous, water-filled aqueous pore directly through the dense lipid bilayer.
The single most famous primary example of this brilliant beta-barrel architecture involves massive proteins called porins. Porins are highly specialized channel-forming proteins that actively utilize this rigid cylindrical beta-barrel structure to effortlessly penetrate the dense cellular membrane. By actively creating a highly stable aqueous channel, these functional proteins enable the rapid, passive passage of vital hydrophilic molecules and essential charged ions. Without these sophisticated, hollow molecular barrels, highly essential hydrophilic nutrients could never successfully cross the tightly packed, strongly hydrophobic barrier. The specialized beta-barrel perfectly demonstrates how biological molecules cleverly fold into unique geometric shapes to completely overcome severe thermodynamic constraints.
12. Plasma Membrane Structure: Lipid-Anchored Proteins
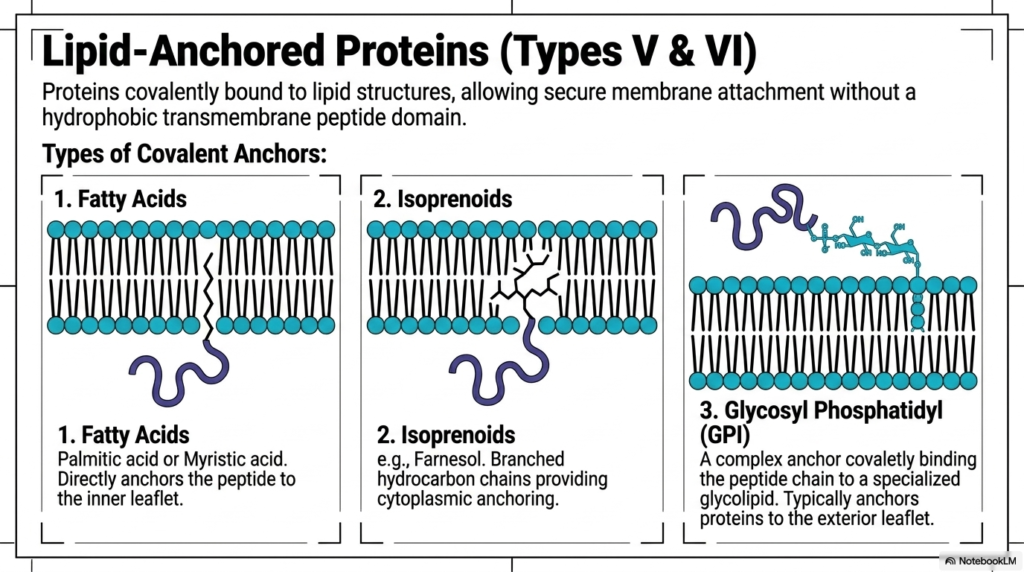
Not all essential boundary proteins require a massive hydrophobic peptide domain to remain securely attached. Within the highly dynamic Plasma Membrane Structure, Types V and VI are strictly represented by vital lipid-anchored proteins. These truly fascinating molecular structures are completely and firmly covalently bound directly to native lipid structures already deeply embedded in the bilayer matrix. This brilliant chemical strategy enables incredibly secure membrane attachment without using a heavy, difficult-to-synthesize hydrophobic transmembrane peptide domain. By specifically using existing structural lipids as permanent molecular anchors, the sophisticated cellular system conserves substantial amounts of essential biosynthetic energy while retaining immense functional versatility.
The biological cell carefully utilizes several completely distinct types of permanent covalent anchors to secure these functional proteins firmly within the Plasma Membrane Structure. The first major structural category relies entirely on specific fatty acids, using either palmitic acid or myristic acid. These notably long hydrocarbon chains directly and permanently anchor the functional peptide chain specifically to the inner cytoplasmic leaflet. Alternatively, the intricate Plasma Membrane Structure actively utilizes complex biological isoprenoids, such as farnesol, for specialized cytoplasmic anchoring. These highly branched hydrocarbon chains forcefully wedge themselves deeply into the dense hydrophobic core, continually providing exceptional mechanical stability for the specifically attached intracellular signaling protein.
The single most complex biochemical anchoring strategy exclusively involves the highly sophisticated Glycosyl Phosphatidylinositol, universally known as the GPI anchor. This massive, highly complex biochemical anchor binds the vital peptide chain directly to a highly specialized, bulky external glycolipid via a strong covalent bond. Unlike the noticeably simpler internal fatty acid or branched isoprenoid anchors, the massive continuous GPI complex typically safely anchors vital functional proteins exclusively to the exterior cellular leaflet. These critical exterior-facing GPI-anchored molecules play an absolutely indispensable, continuous role in advanced enzymatic surface activity and physical cellular adhesion. Together, these incredibly diverse permanent lipid anchors strictly guarantee that every functional biological protein securely reaches its exact required destination within the Plasma Membrane Structure.
13. Plasma Membrane Structure: Synthesizing the Components

A truly comprehensive understanding of advanced cellular biology directly demands carefully synthesizing all the diverse structural elements discussed. The complete, fully functional Plasma Membrane Structure relies heavily on the highly coordinated, continuous interaction of several distinct essential biochemical components. Fundamental phospholipids seamlessly form a continuous five-nanometer amphipathic bilayer core, providing the highly mobile two-dimensional liquid matrix that is critical for all known biological life. Without this primary structural matrix, which cleverly contains distinct components such as vital phosphatidylcholine and specialized sphingomyelin, basic cellular containment would instantly and catastrophically fail. This dynamic, continuously shifting lipid sea powerfully establishes the strict physical rules governing all molecular movement, effectively creating the ultimate biological stage for complex cellular interactions.
Moving noticeably beyond the basic primary matrix, the dynamic Plasma Membrane Structure actively and continuously integrates several highly specialized regulatory and protective structural components. Vital cholesterol acts universally as the absolutely indispensable bidirectional thermodynamic fluidity buffer, intelligently stabilizing highly fluid membranes while aggressively increasing internal fluidity in dangerous frozen semicrystalline states. Simultaneously, highly complex glycolipids and massive glycoproteins constantly combine to completely form the dense, protective exterior glycocalyx hydration layer. The asymmetric Plasma Membrane Structure strictly restricts these highly vital carbohydrate variations, explicitly including bulky gangliosides and complex oligosaccharides, exclusively to the exterior cellular leaflet. This exceptionally precise spatial arrangement continuously ensures the vulnerable cell remains safely physically protected.
Finally, the truly immense functional capabilities of the biological boundary depend entirely on seamlessly integrated, highly sophisticated molecular machinery. Massive integral proteins continuously cross the incredibly dense bilayer via sophisticated hydrophobic right-handed alpha-helices or massive antiparallel beta-barrels to successfully function as vital sensory receptors and structural anchors. Meanwhile, specialized covalent lipid anchors safely secure essential auxiliary functional proteins without ever needlessly penetrating the highly dangerous hydrophobic bilayer core. Crucially, highly specialized translocators, continuously functioning as active flipases, strictly regulate essential membrane asymmetry by forcibly transporting selected phospholipids across the hydrophobic core. Together, these incredibly diverse biochemical components flawlessly construct the ultimate Plasma Membrane Structure, thereby ensuring continued cellular survival and complex physiological function.
Please read our Content Disclaimer Statement.
Check out our social media channels: