74. Membrane Ion Channels: Structure, Function and Mechanisms
Imagine a bustling city without doors, gates, or traffic lights. Chaos would reign. The human body faces a similar challenge at the cellular level. How do cells communicate, trigger muscle contractions, or generate thoughts without letting the outside environment flood in uncontrollably? The answer lies in specialized protein structures. This slide deck explores the fundamental architecture and function of these crucial gatekeepers. By examining their biochemical structures, the presentation reveals the precise mechanisms that enable human life, moving from nanoscale protein folding to systemic physiological actions.
Slide 1: Membrane Ion Channels as Molecular Gatekeepers
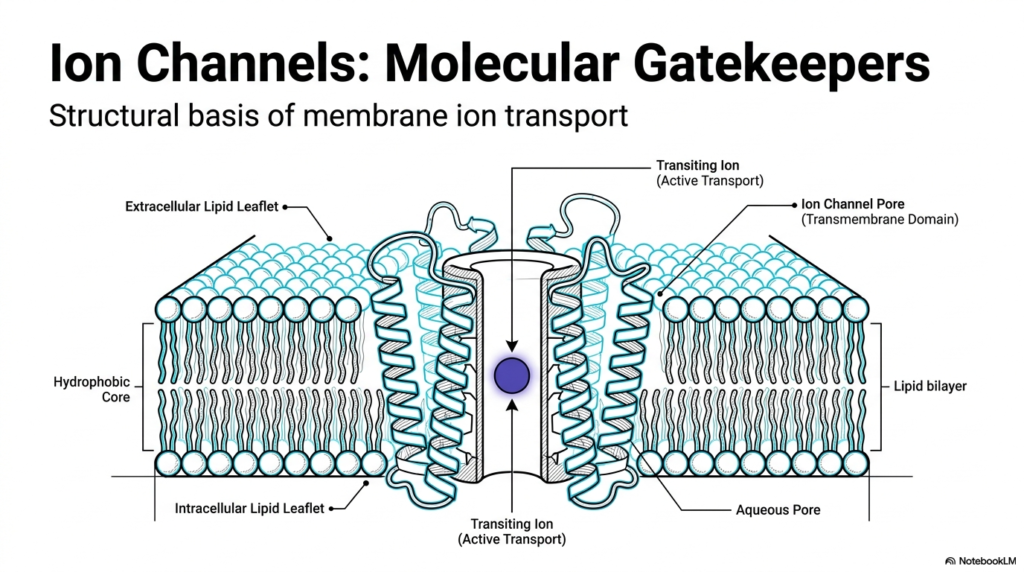
The biological cell is a fortress. Its outer wall, the lipid bilayer, presents a formidable barrier to water-soluble molecules and charged particles. This first image illustrates the foundational concept of Membrane Ion Channels. These structures are not mere holes in the wall; they are sophisticated molecular machines. The diagram showcases the hydrophobic core of the lipid bilayer, which naturally repels charged ions. To overcome this, Membrane Ion Channels provide a highly regulated aqueous pore. This specialized passageway allows transiting ions to cross the membrane safely.
Notice the intricate folding of the protein structures within the lipid environment. The transmembrane domain of Membrane Ion Channels consists of specific amino acid sequences that comfortably anchor the structure within the fatty lipid leaflets. By shielding the charged ions from the hydrophobic core, these proteins facilitate transport. While the slide notes active transport for some transiting ions, the broad family of these channels mostly enables passive, gradient-driven flow. The architecture ensures that only specific materials can pass through the gates.
The structural basis of membrane ion transport relies heavily on the specific arrangement of these protein domains. The extracellular and intracellular lipid leaflets form a tight seal around the channel. This seal guarantees that the only way into or out of the cell is through the designated pore. Students of biochemistry must recognize that this fundamental design is conserved across almost all forms of life. Without this highly selective permeability, cellular homeostasis would instantly fail.
Understanding this basic structural layout provides the necessary groundwork for all subsequent studies in advanced cellular physiology. The biological cell uses these microscopic gateways to continually maintain delicate electrical and chemical balances across its borders. This constant, dynamic regulation serves as the physical basis for everything from basic cellular hydration to complex nerve signaling. Recognizing the sheer physical elegance of this biological solution highlights the extraordinary power of evolutionary biochemistry.
Slide 2: Overcoming the Lipid Barrier with Membrane Ion Channels
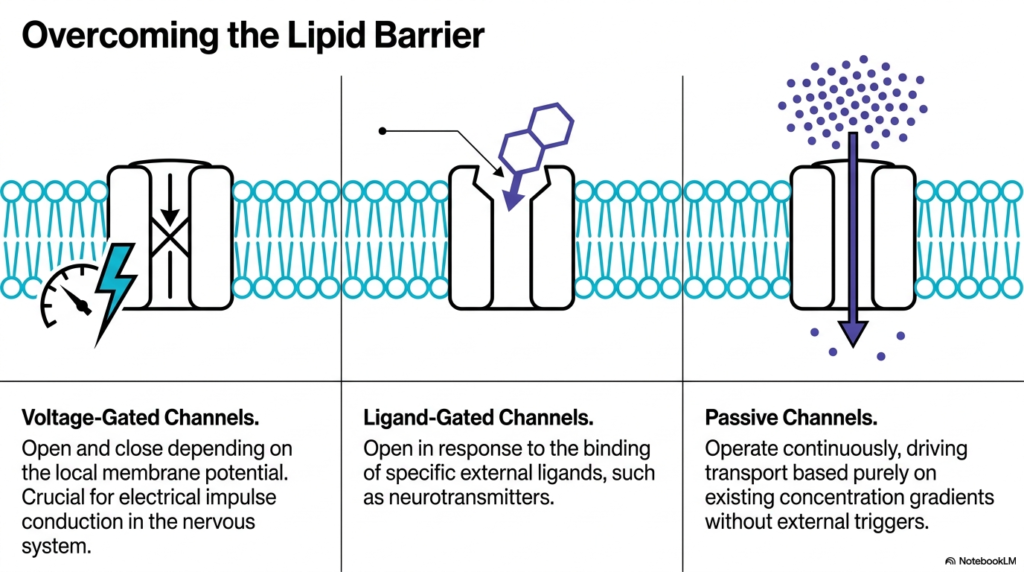
Cells require different types of access controls depending on their immediate physiological needs. This slide categorizes Membrane Ion Channels into three distinct operational classes. The visual breaks down voltage-gated, ligand-gated, and passive channels. Each type solves the problem of the lipid barrier using a completely different biochemical trigger. Understanding these three classifications is essential for grasping how Membrane Ion Channels coordinate complex biological responses across different tissues. The diversity in gating mechanisms allows for extraordinary regulatory control.
Voltage-gated channels act as the cell’s electrical sensors. They open and close in direct response to fluctuations in the local membrane potential. When the electrical charge across the membrane shifts, the protein physically changes shape to allow ion transit. This specific class of Membrane Ion Channels is the absolute foundation for electrical impulse conduction. Without them, the nervous system could not transmit signals, rendering the entire brain and peripheral nervous system entirely nonfunctional. Students must appreciate that this voltage sensitivity represents a marvel of molecular engineering.
Ligand-gated channels operate via chemical recognition. Instead of sensing electricity, they wait for specific external molecules, known as ligands, to bind to their receptor sites. Neurotransmitters often serve as these chemical keys. Once the neurotransmitter binds, the channel unlocks. This allows target cells to translate a chemical message from a neighboring cell into an immediate electrical or biochemical response. This mechanism is primarily how neurons communicate with muscles and other nerves.
Passive channels, often called leak channels, operate quite differently from the other two active types. They remain continuously open, providing a steady, unregulated pathway for ions to move. This movement relies entirely on existing concentration gradients without requiring an external electrical or chemical trigger. These channels are fundamental to establishing the cell’s baseline resting membrane potential. Together, these three distinct channel types perfectly orchestrate cellular life.
Slide 3: Membrane Ion Channels Driving the Neurological Action Potential
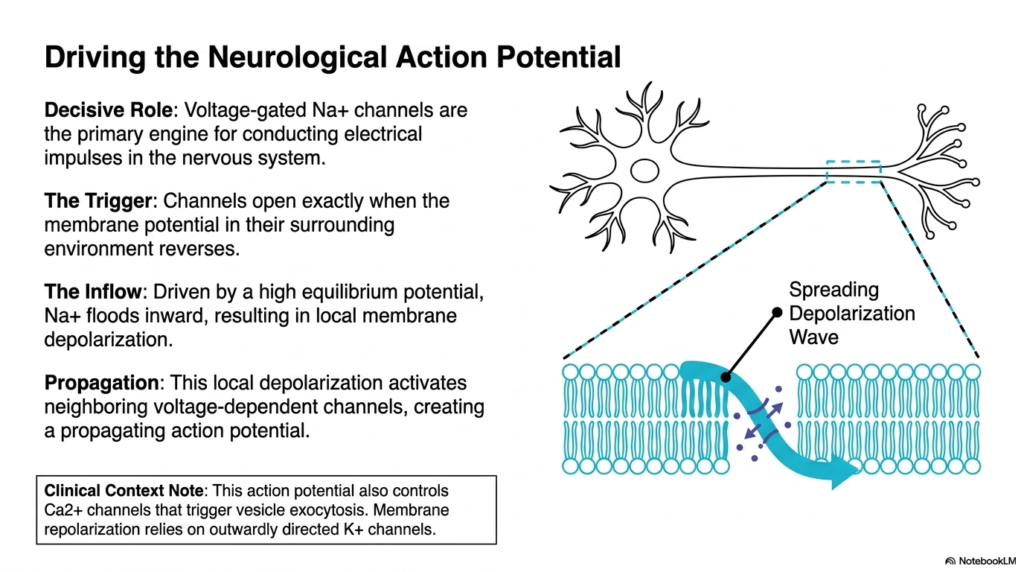
The nervous system relies on rapid communication to function. This slide details how specific Membrane Ion Channels act as the primary engines for this electrical signaling. The diagram highlights a neuron and the spreading wave of depolarization along its membrane. The decisive role belongs to voltage-gated sodium channels. These specialized Membrane Ion Channels drive the massive influx of positive charge needed to fire a nerve impulse. They are the biochemical catalysts for every thought and movement.
The entire process begins with a precise trigger event. The sodium channels remain firmly closed until the membrane potential in their surrounding environment reverses. Once this threshold is crossed, the channels snap open instantly. Because the cellular environment maintains a high equilibrium potential, sodium ions possess a strong driving force to enter the cell. This sudden, massive inflow of positively charged sodium completely flips the local electrical state, causing localized depolarization.
This localized depolarization does not remain confined to one spot. The influx of positive charge alters the membrane potential of the adjacent cellular membrane. This local change activates neighboring voltage-dependent channels in a chain reaction. The result is a self-propagating action potential that races down the length of the nerve fiber. The study of Membrane Ion Channels reveals how local atomic shifts scale up to create macroscopic nerve conduction spanning long distances.
The clinical context of this action potential is profoundly important. The traveling depolarization wave eventually reaches the nerve terminal, where it controls calcium channels that trigger vesicle exocytosis. This allows chemical messages to cross synapses. Furthermore, the cell must reset its electrical state after firing. This critical repolarization phase relies on outwardly directed potassium channels. Therefore, a perfectly timed sequence of different channel activations is required for a single, complete nerve impulse cycle.
Slide 4: Membrane Ion Channels and Massive Peptide Architecture
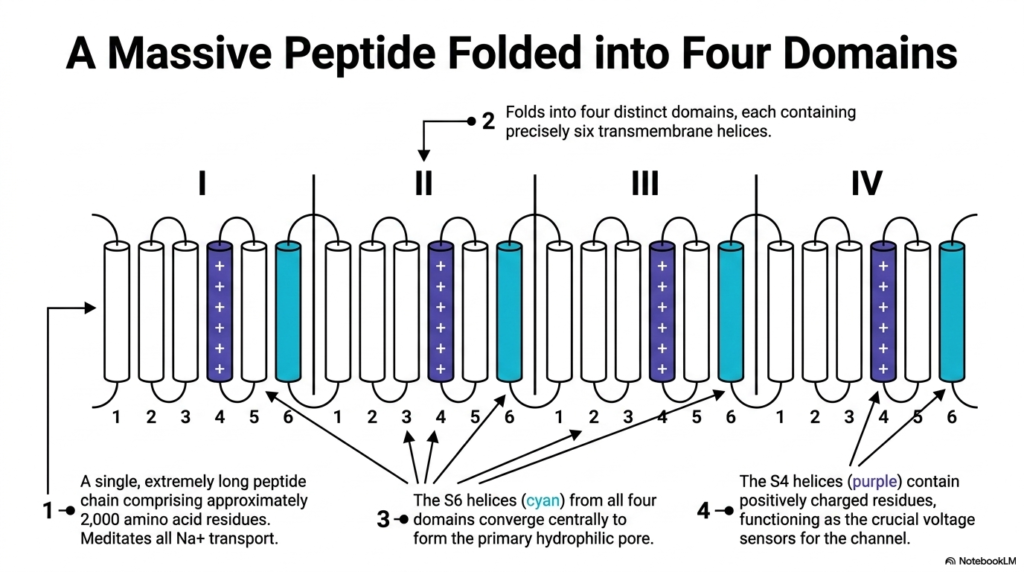
To understand how voltage-gated mechanisms function, one must examine the massive protein architecture involved. This slide illustrates the complex topology of voltage-gated sodium Membrane Ion Channels. The graphic depicts a single, incredibly long peptide chain comprising approximately 2,000 amino acid residues. This continuous chain folds intricately to mediate all cellular sodium transport. The sheer size and complexity of these Membrane Ion Channels highlight the evolutionary importance of precise electrical regulation.
The structure is highly organized, folding into four distinct, repeating domains labeled 1 through 4. Within each of these four domains, the protein crosses the cell membrane exactly six times. These transmembrane crossings form six distinct alpha-helices per domain. By arranging these twenty-four helices in a circular pattern, the massive protein creates a highly functional three-dimensional shape. This repeating motif is a classic biochemical signature of many critical voltage-responsive channels.
Two specific types of helices perform the most critical tasks. The S6 helices, colored cyan in the diagram, converge at the center from all four domains. Together, they create the primary hydrophilic pore through which the sodium ions actually travel. In contrast, the S4 helices, shown in purple, play a very different role. The S4 helices contain positively charged amino acid residues. These specific regions function as the crucial voltage sensors for the entire structure.
The division of labor within these Membrane Ion Channels is remarkable. The structure dedicates specific physical domains exclusively to environmental sensing, while other distinct regions constitute the actual transport machinery. This brilliant modular design allows the massive protein complex to respond instantly to electrical changes without compromising the structural integrity of the central ion pore. Such detailed structural and architectural knowledge is absolutely crucial for modern pharmacology. It aids in the development of targeted neurotoxins and therapeutic local anesthetics that bind directly to these highly specific functional domains.
Slide 5: The Mechanical Spring of Membrane Ion Channels
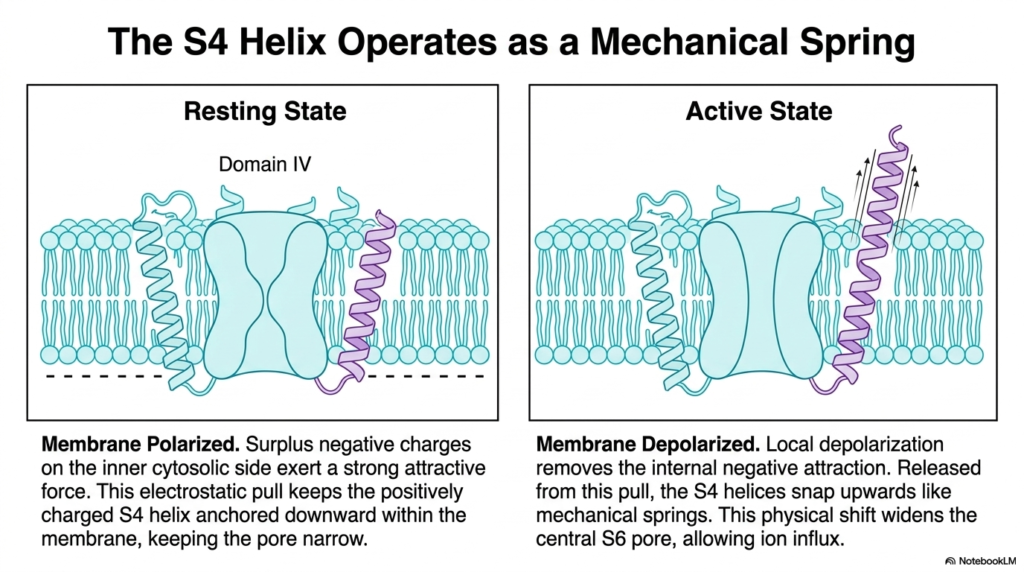
The abstract concept of voltage sensing becomes clear when viewed as a mechanical process. This slide demonstrates how the S4 helix in certain Membrane Ion Channels functions as a mechanical spring. The diagram contrasts the resting state of the channel with its active state. By examining Domain IV, students can visualize the exact physical movement that opens the pore. This translation of electrical potential into mechanical movement is a hallmark of Membrane Ion Channels.
During the resting state, the cellular membrane remains strongly polarized. A large surplus of negative electrical charges lines the inner cytosolic side of the cell membrane. Because the S4 helix contains positively charged amino acid residues, this internal negativity exerts a strong attractive electrostatic force. This constant downward pull keeps the S4 helix firmly anchored deeply within the membrane layer. In this lowered position, the channel remains locked, keeping the central pore extremely narrow.
Activation occurs when the membrane undergoes depolarization. As the local area loses its negative internal charge, the strong attractive force vanishes. Freed from this electrostatic pull, the positively charged S4 helices violently snap upwards toward the extracellular space, mimicking the release of a coiled mechanical spring. This dramatic upward physical shift tugs on the surrounding protein structure. The movement directly pulls the central S6 pore open, creating a wide channel for rapid sodium influx.
This fascinating mechanism proves that Membrane Ion Channels are highly dynamic molecular machines. The cellular membrane potential does not merely shift invisible electrical fields; it causes massive, rapid physical movements in solid protein structures. This precise electromechanical coupling occurs in fractions of a millisecond. Understanding this spring-loaded mechanism provides a tangible, physical explanation for how nerve cells can fire so incredibly quickly. Medical students studying neurological arrhythmias often trace systemic diseases to minute genetic mutations that affect these very molecular springs.
Slide 6: Chemical Triggers for Membrane Ion Channels
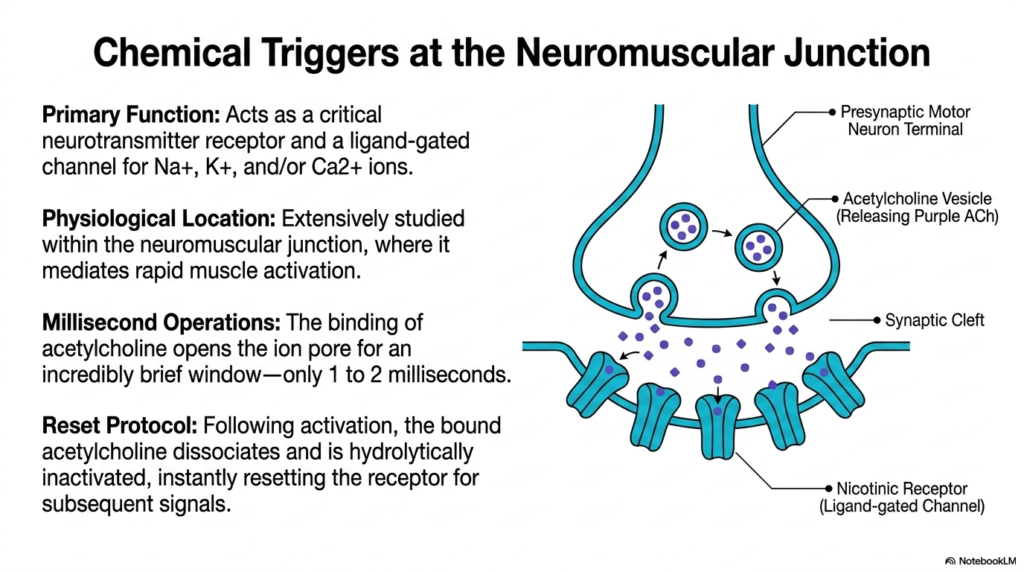
While voltage triggers are vital, chemical communication is equally crucial for human physiology. This slide focuses on chemical triggers at the neuromuscular junction and illustrates a different class of Membrane Ion Channels. The diagram depicts a presynaptic motor neuron terminal releasing purple acetylcholine molecules into the synaptic cleft. These molecules target the nicotinic receptor, a classic ligand-gated channel. This specific class of Membrane Ion Channels translates chemical messages into rapid muscle activation. Studying this junction provides the perfect biochemical model for understanding synaptic transmission throughout the body.
The primary function of the nicotinic receptor is to act as a critical neurotransmitter receptor. Once activated, it serves as a non-selective ligand-gated channel, allowing sodium, potassium, and sometimes calcium ions to flow across the membrane. This receptor is extensively studied due to its physiological location within the neuromuscular junction. Whenever the brain commands a muscle to move, it relies entirely on these specific receptors to bridge the gap between nerve and muscle tissue.
The speed of this chemical transmission is difficult to overstate. The binding of acetylcholine opens the ion pore for an incredibly brief window of time, typically lasting only one to two milliseconds. This millisecond operation permits a massive, nearly instantaneous influx of ions that sparks muscle contraction. The sheer speed of these Membrane Ion Channels enables humans to perform highly coordinated, rapid physical movements such as typing or sprinting.
For the system to function correctly, the signal must terminate just as quickly as it begins. Following activation, a strict reset protocol initiates. The bound acetylcholine molecules rapidly dissociate from the receptor. Enzymes in the synapse then hydrolytically inactivate the neurotransmitter. This instant clearance resets the receptor, ensuring the channel closes promptly. Without this rapid clearance, muscles would remain in a state of continuous, rigid paralysis, unable to relax in response to subsequent functional signals.
Slide 7: Precision Assembly of Pentameric Membrane Ion Channels
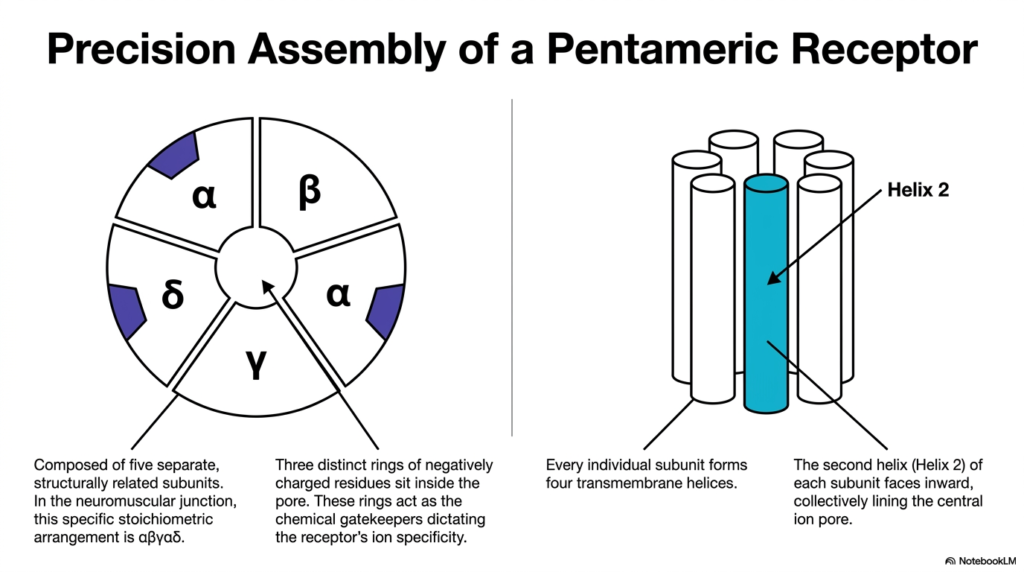
The functional speed of chemical receptors requires precise structural engineering. This slide details the precision assembly of the nicotinic acetylcholine receptor, a prominent member of the ligand-gated Membrane Ion Channels. The graphic breaks down the complex pentameric structure. In the neuromuscular junction, this specific stoichiometric arrangement consists of two alpha subunits, one beta, one delta, and one gamma subunit. This precise five-part construction is a hallmark of many critical Membrane Ion Channels.
The complex includes structural redundancies and highly specialized zones. Every individual subunit within this pentamer forms four distinct transmembrane helices. When these five subunits assemble in a ring, they create a functional pore. The diagram shows that the second helix (Helix 2) of each subunit faces inward. Together, these five inward-facing helices collectively line the central ion pore, forming the actual physical boundary that interacting ions must navigate.
The architecture does more than just form a simple tube. Three distinct rings of negatively charged amino acid residues sit strategically inside the pore. These carefully placed negative charges act as the chemical gatekeepers for the receptor. They strongly attract positively charged cations, such as sodium and potassium, while aggressively repelling negatively charged anions. This strategic placement of electrical charge dictates the receptor’s overall ion specificity with remarkable biochemical precision. Without these charged rings, the channel would lose its selective filtering capability, leading to catastrophic cellular depolarization and systemic physiological failure.
This level of architectural detail is vital for understanding pharmacological targeting. Many toxins, venoms, and pharmaceutical drugs operate by binding to specific interfaces between these precise subunits. By interacting with the alpha or delta components of these Membrane Ion Channels, external chemicals can block or forcefully open the central pore. Therefore, mastering this pentameric subunit geometry provides medical students with the foundational knowledge needed to understand neuropharmacology and advanced anesthesiology.
Slide 8: Membrane Ion Channels and Ligand-Induced Expansion
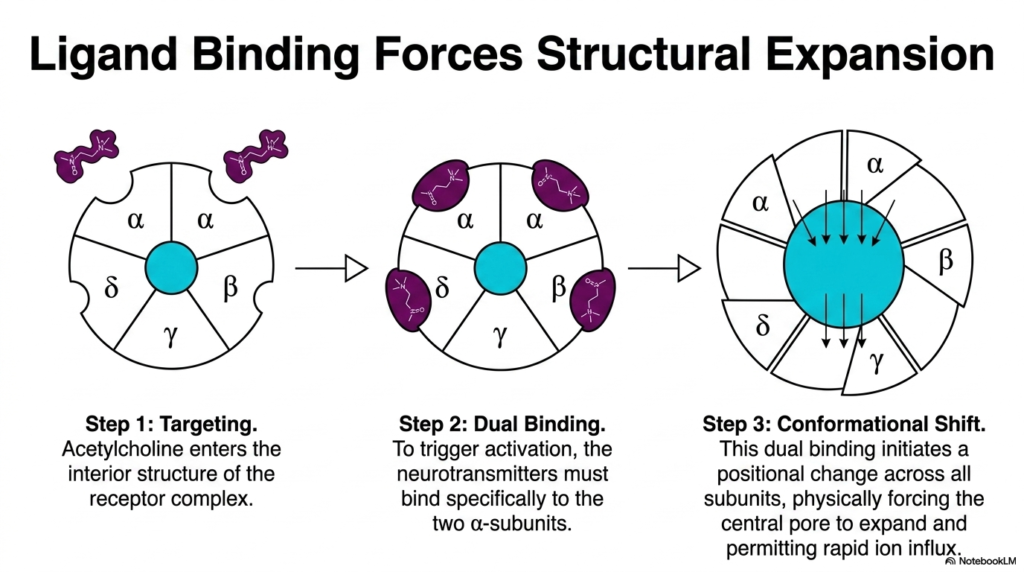
Structure dictates function, and binding dictates movement. This slide illustrates exactly how acetylcholine binding physically opens the nicotinic receptor. It provides a three-step visual guide to the structural expansion of these ligand-gated Membrane Ion Channels. The process highlights the incredible mechanical shifts that occur at the atomic level. This visual sequence clearly demonstrates how Membrane Ion Channels act as dynamic transformers, converting chemical binding energy into significant physical pore dilation.
The mechanism begins with the targeting phase. Acetylcholine molecules, released from the adjacent nerve terminal, enter the interior structure of the receptor complex. The specific geometry of the receptor guides these chemical messengers directly toward their intended binding sites. However, a single molecule is insufficient to trigger the complete opening mechanism. To trigger full activation, the receptor demands dual binding. The neurotransmitters must bind to both alpha subunits simultaneously. This simultaneous engagement is an elegant evolutionary design ensuring strict physiological control over muscle contraction.
This dual binding requirement serves as a biological safety mechanism. It ensures that the channel only opens in response to a concentrated, intentional release of neurotransmitters, preventing accidental firing from stray background molecules. Once both alpha subunits bind an acetylcholine molecule, the final conformational shift occurs. This dual binding initiates a cascade of positional changes across all five subunits. The protein structure literally twists and expands under the new chemical tension.
This coordinated shift physically forces the central pore to expand outward. The inward-facing helices rotate out of the way, removing the physical blockade. This expansion instantly enables the rapid influx of ions necessary for cellular signaling. The study of these specific Membrane Ion Channels reveals that ligand gating is not a magical unlocking process. Instead, it relies on forceful, mechanical leverage exerted by tiny chemical messengers on massive protein structures.
Slide 9: The First High-Resolution Blueprint of Membrane Ion Channels
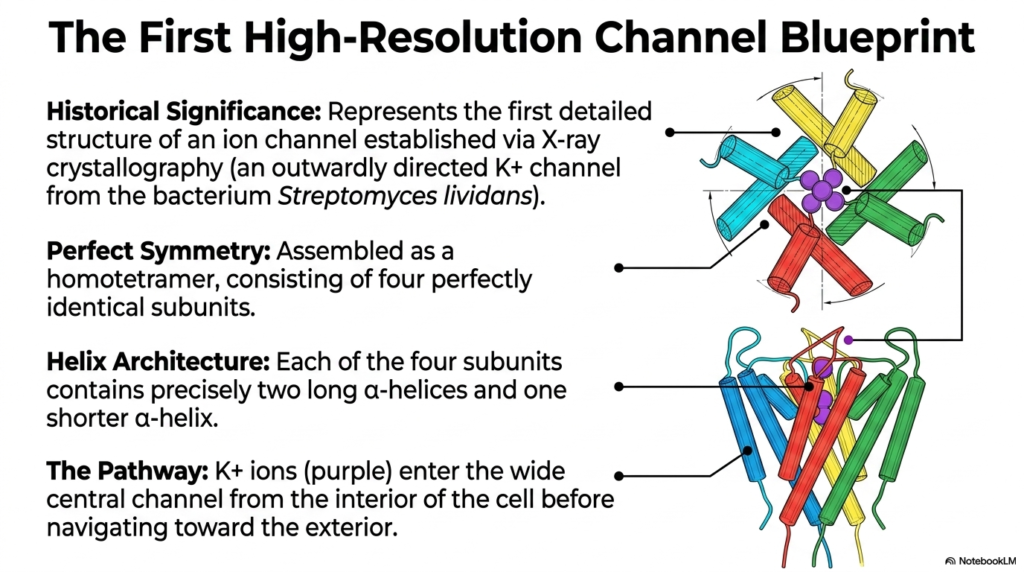
The transition from physiological theory to structural certainty required a massive technological breakthrough. This slide details the historical significance of the very first high-resolution channel blueprint. Researchers used X-ray crystallography to determine the exact atomic structure of an outward-facing potassium channel from the bacterium Streptomyces lividans. This monumental achievement completely revolutionized our understanding of Membrane Ion Channels. Finally, scientists could visually confirm the long-theorized mechanisms of Membrane Ion Channels at the atomic level.
The X-ray structure revealed a masterclass in biological symmetry. The bacterial potassium channel is assembled as a homotetramer, meaning it consists of four perfectly identical subunits. This simple but elegant design contrasts with the complex heteromeric structure of the nicotinic receptor. The helix architecture is equally streamlined. Each of the four identical subunits contains precisely two long alpha-helices and one shorter, angled alpha-helix. Together, these elements frame the central conduction pathway.
The visual model traces the exact pathway ions take during transport. Potassium ions enter the wide central cavity of the channel from the interior cytosolic environment of the cell. The structure acts like a broad funnel, easily gathering the positively charged ions and directing them toward the narrowest part of the pore. Before navigating toward the exterior, the ions must align perfectly within this central corridor. This wide vestibule lowers the energy barrier for ions, allowing them to remain fully hydrated in an aqueous pool before they encounter the highly restrictive filtering segment of the protein.
The structural conservation seen in this bacterial model applies directly to human biochemistry. The basic homotetrameric architecture discovered in Streptomyces lividans is widely shared among human voltage-gated potassium channels. By studying this primitive blueprint, medical researchers gained profound insights into human cardiac rhythm and neurological repolarization. This specific discovery demonstrated that the fundamental engineering of Membrane Ion Channels was perfected very early in biological history and has been preserved across billions of years of evolution.
Slide 10: Atomic Geometry in Membrane Ion Channels
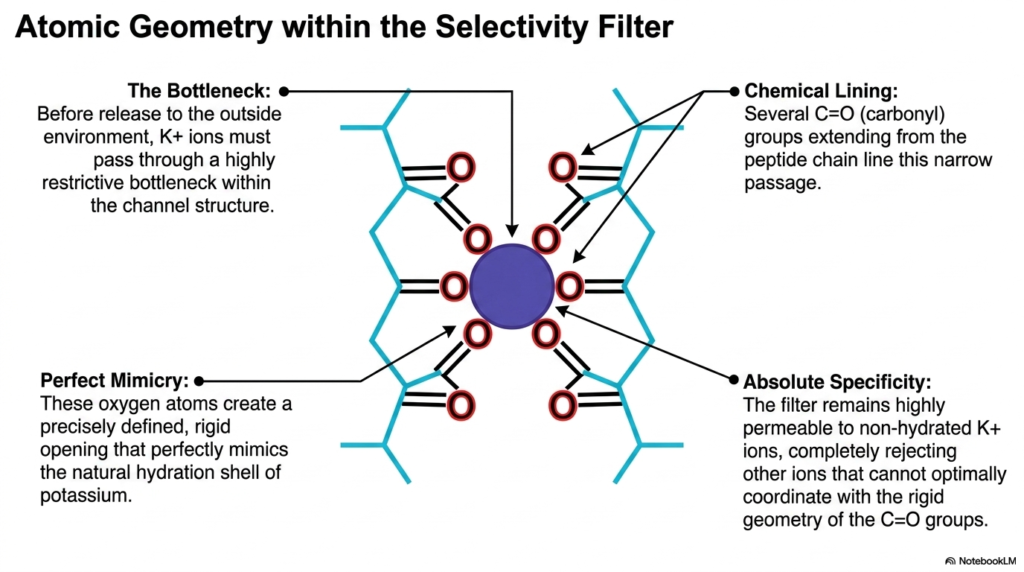
The most perplexing question in channel biology is how a large pore can be highly specific. This slide examines the atomic geometry within the selectivity filter of potassium channels. This specific region solves the mystery of absolute ion specificity. Before release to the outside environment, potassium ions must pass through a highly restrictive bottleneck within the channel structure. This segment of the membrane ion channels represents one of the most precisely engineered spaces in biology. Understanding this tiny selectivity filter reveals how cells maintain their crucial internal chemistry without mixing vital electrolytes.
The chemical lining of this bottleneck provides the mechanism of selection. Several carbonyl groups, representing carbon double-bonded to oxygen, extend directly from the surrounding peptide chain. These groups line the narrow passage, pointing their negatively charged oxygen atoms inward. These oxygen atoms create a precisely defined, rigid opening. This specific geometry achieves perfect mimicry. The spacing of the oxygen atoms perfectly mimics the natural hydration shell of a dissolved potassium ion.
When a potassium ion enters this filter, it sheds its surrounding water molecules. The rigid carbonyl oxygen atoms seamlessly replace the water, stabilizing the ion energetically. The filter remains highly permeable to these non-hydrated potassium ions. They glide through the passage with remarkable speed and efficiency. However, the system completely rejects other ions, such as sodium. Even though sodium is smaller in size, it is structurally incompatible with this rigidly designed filter.
Sodium cannot optimally coordinate with the fixed geometry of the carbonyl groups. Because it cannot perfectly bind to all the oxygen atoms, it cannot comfortably shed its water shell. Therefore, the energetic cost of entering the filter is simply too high. This absolute specificity prevents chaotic ion crossover. It ensures that these specific Membrane Ion Channels transport only potassium, maintaining the strict electrical gradients required for healthy cellular function and repolarization.
Slide 11: Architectural Comparison of Membrane Ion Channels
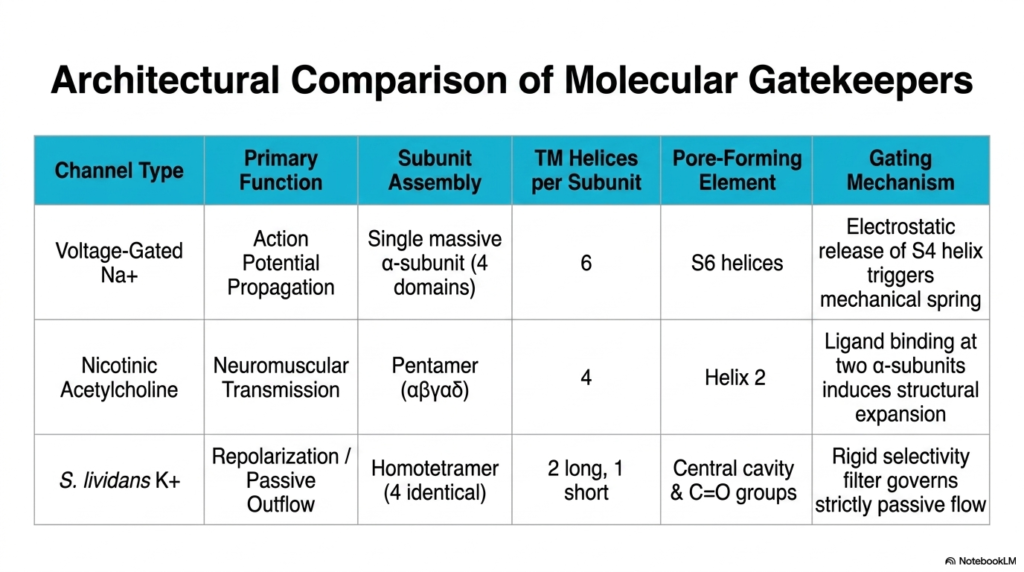
Synthesizing structural knowledge is vital for a comprehensive biochemical understanding. This slide provides an architectural comparison of the previously discussed molecular gatekeepers. The comparative table perfectly summarizes the diverse structural strategies employed by different Membrane Ion Channels. By placing the voltage-gated sodium channel, the nicotinic acetylcholine receptor, and the bacterial potassium channel side by side, the major differences in biological design become immediately apparent. The vast diversity of Membrane Ion Channels is a testament to biological adaptation.
The table highlights the primary function and subunit assembly of each channel type. The sodium channel mediates action potential propagation using a single massive alpha-subunit folded into four domains. In stark contrast, the nicotinic receptor drives neuromuscular transmission using a pentameric assembly of five distinct subunits. Meanwhile, the simpler bacterial potassium channel mediates passive outflow via a homotetramer of four identical subunits. These distinct physical assemblies correlate directly with their specialized biological roles. Understanding these variations helps explain why certain pharmaceutical drugs affect only specific tissues, targeting one subunit assembly while ignoring others entirely.
The pore-forming elements and transmembrane helices also vary significantly. The complex sodium channel requires twenty-four helices in total, using the S6 helices for the pore. The pentameric acetylcholine receptor relies on 20 helices, with Helix 2 lining the pathway. The streamlined potassium channel uses only twelve helices total, relying on a central cavity and carbonyl groups. Despite these differences, they all successfully create secure, functional pathways across the hydrophobic lipid bilayer.
Finally, the table contrasts their gating mechanisms. The sodium channel utilizes the electrostatic release of the S4 helix to trigger a mechanical spring. The nicotinic receptor relies on dual ligand binding to induce a massive structural expansion. In this specific model, the potassium channel uses a rigid selectivity filter to govern strictly passive flow. Comparing these mechanisms helps students appreciate how diverse Membrane Ion Channels regulate every aspect of dynamic cellular physiology.
Slide 12: Core Biochemical Truths of Membrane Ion Channels

We conclude this comprehensive academic review by summarizing the most critical takeaways. This final slide presents the core biochemical truths for the practitioner. It distills the complex structural data into three fundamental principles that govern all Membrane Ion Channels. These principles bridge the gap between abstract molecular geometry and tangible clinical outcomes. For medical students and future researchers, mastering these three foundational concepts of membrane ion channels is an absolute necessity for clinical practice. Without internalizing these rules, one cannot fully comprehend how specialized neurotoxins, local anesthetics, or targeted cardiac medications actually interact with the human body at the foundational molecular level.
The first truth emphasizes that mechanical action is inherently electro-chemical. Membrane potentials do not simply shift abstract electrical charges around the cell. Instead, they cause massive, rapid physical movements in solid protein structures. The S4 helix acting as a loaded molecular spring perfectly exemplifies this. When diagnosing neurological disorders or cardiac arrhythmias, practitioners must remember they are often dealing with microscopic physical failures of these moving protein parts, not just electrical imbalances.
The second truth states that geometry dictates absolute specificity. Whether using distinct rings of negative charge, as in the nicotinic receptor, or precisely angled carbonyl groups that mimic hydration shells in potassium channels, the architecture is flawlessly deliberate. This rigid pore architecture prevents chaotic ion crossover. The exact sub-angstrom measurements of these channels determine which molecules live or die within the cell. This rigid geometric control is the ultimate basis of all selective biological permeability.
The final truth is that systemic impact stems directly from nano-scale events. The macroscopic propagation of a neurological action potential, or the rapid, powerful contraction of a skeletal muscle, relies entirely on these sub-millisecond conformational shifts. The entire human experience—every heartbeat, breath, and thought—is the sum of billions of Membrane Ion Channels opening and closing in perfectly orchestrated sequence. Understanding this connection allows practitioners to appreciate the profound power of targeted molecular medicine.
Please read our Content Disclaimer Statement.
Check out our social media channels: