78. Protein Synthesis and Maturation: A Biochemical Overview
Imagine a bustling factory where raw materials are transformed into sophisticated machines. This slide explores the core biochemical pathways occurring in the endoplasmic reticulum, detailing how cells manufacture essential biological components. The fundamental goal of this slide deck is to demystify the journey of a nascent polypeptide chain, transforming an abstract concept into a clear, visual roadmap for medical and college students. Understanding these pathways is crucial for mastering cellular biology.
Slide 1: The Foundation of Protein Synthesis: How Cells Build Life
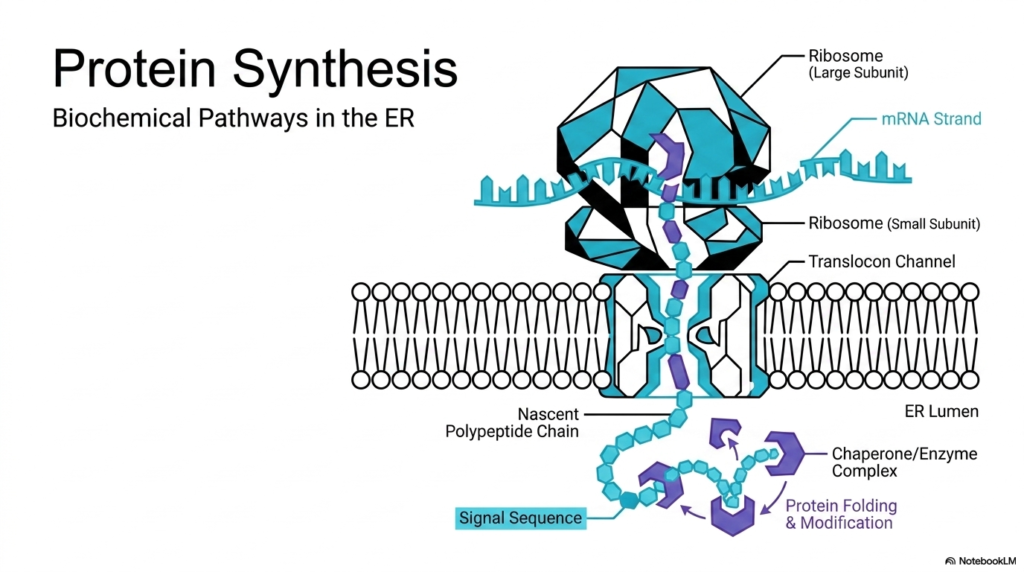
The illustration highlights the early stages of protein synthesis, a highly coordinated process that begins with the ribosome reading an mRNA strand. The ribosome, composed of a large and small subunit, acts as the primary manufacturing plant. As it decodes the genetic instructions, it extrudes a nascent polypeptide chain. This emerging chain features a crucial signal sequence that acts as a molecular targeting code. This sequence directs the entire complex to the endoplasmic reticulum membrane. For students studying cell biology, visualizing this docking process is essential to grasp how the cell compartmentalizes its operations and separates cytosolic functions from secretory pathways.
Once docked at the membrane, the growing chain enters a specialized channel known as the translocon. The translocon serves as a secure gateway, allowing the delicate new molecule to pass safely into the protective environment of the internal organelle lumen. This step is a defining moment in protein synthesis, shifting the location of production from the open cytoplasm to a controlled internal chamber. Inside the lumen, chaperone enzymes wait to assist with folding and structural modifications. These chaperones ensure the linear chain achieves its correct three-dimensional shape, preventing misfolding that could lead to cellular dysfunction or disease.
Without correct folding, the products of protein synthesis cannot perform their intended biological roles. The collaboration between the ribosome, the translocon, and luminal chaperones represents a masterpiece of biochemical engineering. Every component works in perfect harmony to produce functional molecules ready for further transport or immediate use. Grasping these fundamental interactions prepares students to tackle more complex topics like post-translational modifications, glycosylation, and targeted transport in subsequent lessons.
Slide 2: The Journey of Protein Synthesis: Pathways of Secretory Proteins
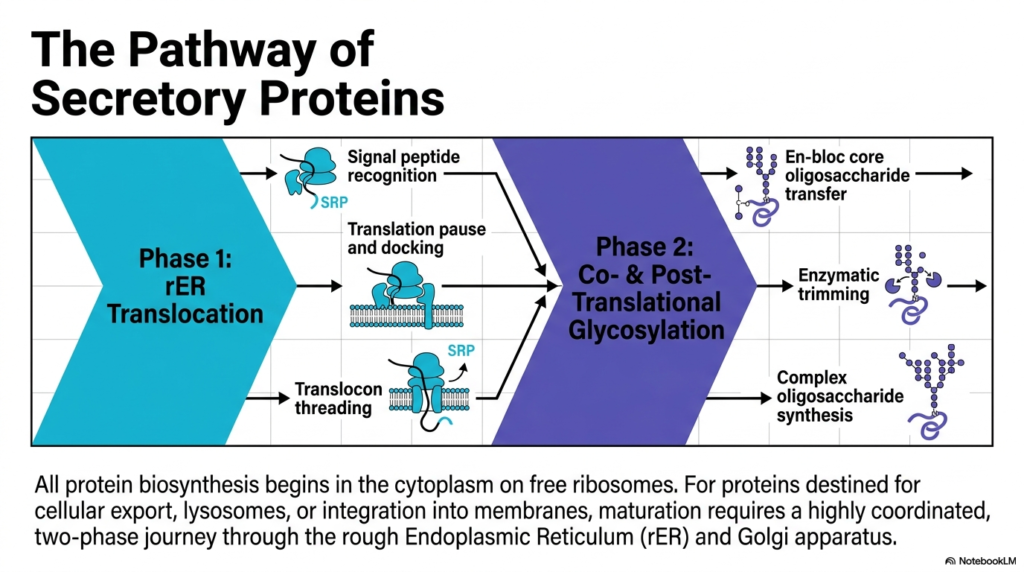
Have you ever tracked a package from a warehouse to your front door? Cells use a similar tracking and delivery system for their molecular products. This slide outlines the two-phase journey that secretory proteins take to reach their final destination. The core purpose of this presentation is to map the specific checkpoints these molecules pass through, providing a clear overview of maturation for future health professionals.
The protein synthesis roadmap begins in the cytoplasm on free ribosomes. However, molecules destined for cellular export, lysosomes, or membrane integration require special handling. This handling involves a coordinated journey through the rough endoplasmic reticulum and the Golgi apparatus. Phase one involves translocation into the lipid boundary. This initial phase requires the recognition of a specific signal peptide by a signal recognition particle. Once recognized, translation pauses, allowing the entire molecular complex to safely dock at the membrane before threading through the translocon channel.
The second phase introduces co-translational and post-translational glycosylation, a critical modification step in protein synthesis. Once the growing chain enters the lumen, a massive carbohydrate structure called a core oligosaccharide transfers en bloc to the molecule. This bulky addition is not the final product. Enzymes immediately begin trimming the sugar structure, refining it for the next stage of transport. This precise editing process prepares the molecule for its eventual departure from the current cellular compartment.
Following the initial enzymatic trimming, the modified molecule travels to the Golgi apparatus, where it undergoes complex oligosaccharide synthesis. This final polishing step ensures the molecular package has the correct biological tags for its specific destination, such as the cell surface or a digestive lysosome. The entire cascade, from the initial pause in protein synthesis to the intricate carbohydrate modifications, demonstrates the immense precision of cellular machinery. For students aiming to understand genetic disorders or metabolic diseases, recognizing how these pathways function in a healthy cell is absolutely fundamental.
Slide 3: Directing Protein Synthesis: The ER Signal Peptide
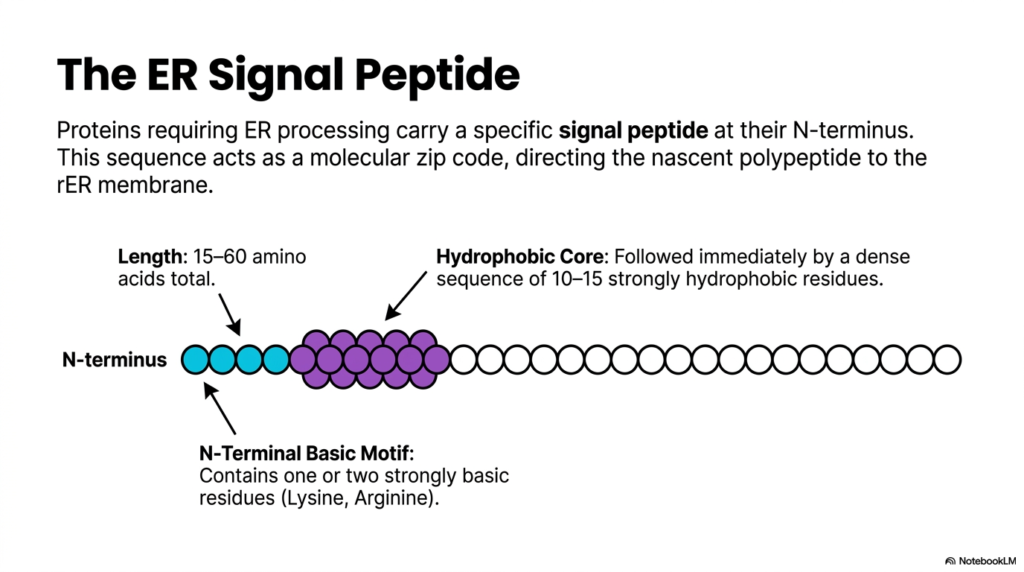
Just as every piece of mail needs a precise zip code to reach its destination, cellular products require clear delivery instructions. This slide details the structure of the ER signal peptide, a sequence that guides molecules to the correct cellular compartment. The fundamental purpose of this slide deck is to break down these targeting mechanisms, showing biology students exactly how amino acid sequences dictate molecular destiny.
Not all molecules produced during protein synthesis enter the secretory pathway. Those that require processing in the endoplasmic reticulum carry a specific signal peptide located at their N-terminus. This sequence typically ranges from 15 to 60 amino acids in total length. The signal peptide acts as a dedicated molecular zip code, ensuring that the nascent polypeptide immediately directs itself to the rough organelle membrane. Without this crucial identifying sequence, the molecule would remain stranded in the cytoplasm, unable to undergo vital maturation steps. The discovery of this zip code system revolutionized our understanding of cell biology.
A closer look at the signal peptide reveals a distinct and highly conserved anatomical structure. At the very beginning of the N-terminus, there is a basic motif containing one or two strongly basic amino acid residues, usually lysine or arginine. These positively charged residues play a vital role in the initial orientation and interaction with the membrane targeting machinery. The presence of these specific amino acids highlights the incredible specificity required for successful protein synthesis and transport.
Immediately following the basic motif lies the hydrophobic core of the signal peptide. This core consists of a dense sequence of 10 to 15 strongly hydrophobic residues. Because the cellular environment is largely aqueous, these water-repelling amino acids naturally seek the lipid-rich environment of the cellular membrane. This physical property drives the initial insertion of the molecule into the translocation channel. By mastering the structure of the signal peptide, students gain profound insight into how simple chemical properties govern complex biological routing during protein synthesis.
Slide 4: Navigating Protein Synthesis: Signal Recognition Particle Dynamics
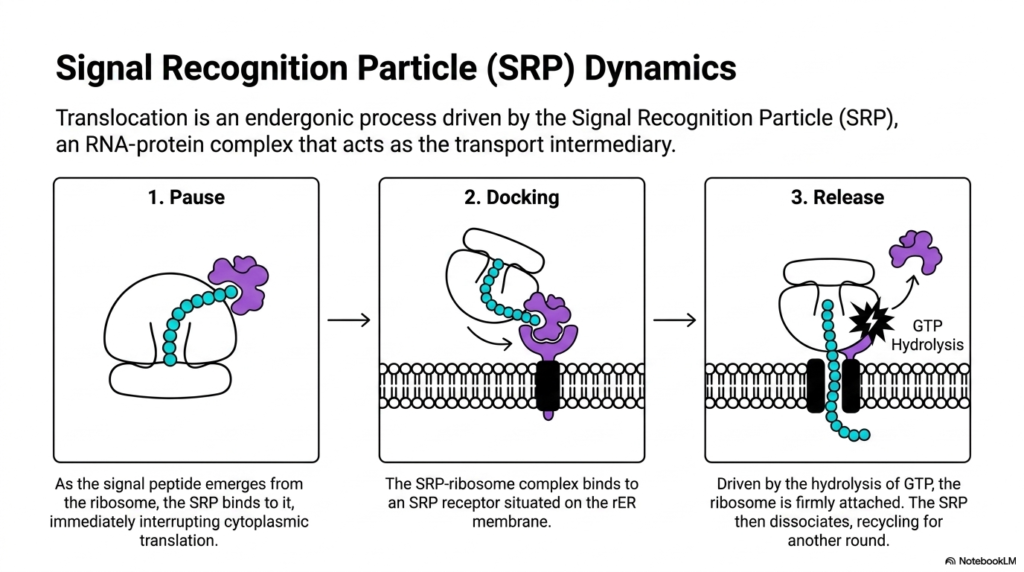
Imagine a tugboat guiding a massive cargo ship safely into a harbor. The cell uses a similar chaperone to guide translating ribosomes to the membrane. This slide examines the dynamics of the Signal Recognition Particle, an essential transport intermediary. The purpose of this presentation is to detail the precise endergonic mechanisms that enable a massive molecular complex to dock with high precision on the cellular membrane.
Translocation across the membrane is not a spontaneous event; it is a highly controlled, endergonic process driven by the Signal Recognition Particle. This particle is an intricate RNA-protein complex that acts as a vital bridge between the cytoplasm and the endoplasmic reticulum. As the signal peptide emerges from the ribosome during the early stages of protein synthesis, the particle recognizes and binds to it. This binding event triggers an immediate pause in cytoplasmic translation. This temporary pause is a brilliant evolutionary adaptation, ensuring the delicate polypeptide remains unfolded until it reaches the safety of the membrane channel.
With translation halted, the second step of the dynamic sequence begins: docking. The combined complex of the particle, the ribosome, and the nascent chain travels to the membrane. Here, it binds to a specific receptor situated on the lipid surface. This docking mechanism ensures that the ribosome aligns perfectly with the translocation channel. The precision of this alignment is a critical quality control checkpoint in protein synthesis, guaranteeing that the growing chain will pass directly into the lumen rather than spilling freely into the cytosol.
The final stage of this interaction involves the release of the transport complex. Driven by the energy released from GTP hydrolysis, the ribosome becomes firmly attached to the membrane channel. Once the connection is secure, the transport particle dissociates from the receptor and the signal peptide. It is then recycled back into the cytoplasm to participate in another round of protein synthesis. Understanding this GTP-dependent cycle allows medical students to appreciate the energy costs and mechanical forces required to build and organize cellular structures.
Slide 5: Advancing Protein Synthesis: Translocon Threading and Cleavage
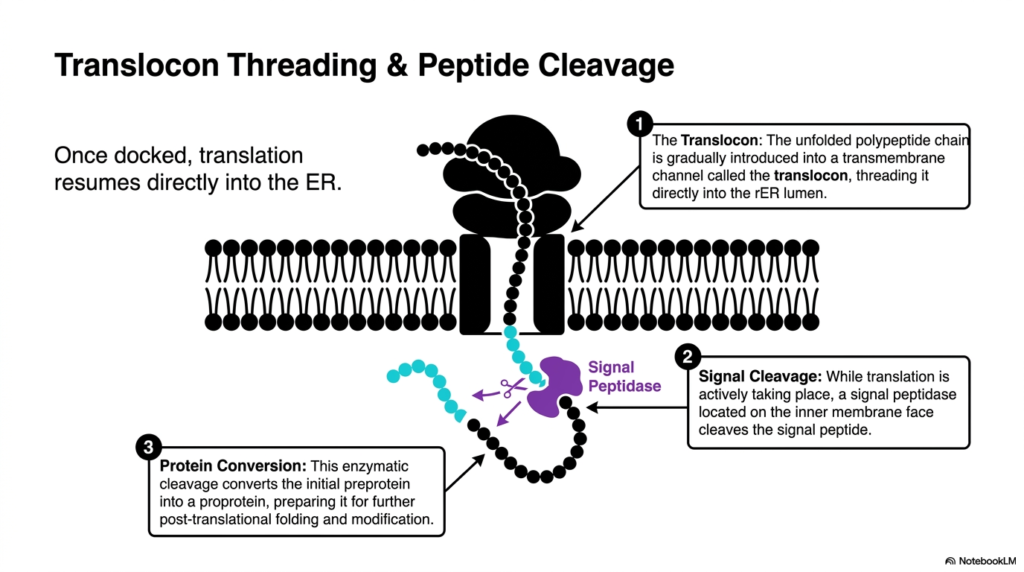
Think of a thread passing through the eye of a needle while simultaneously being trimmed to the perfect length. This slide illustrates the exact moment a growing polypeptide enters the membrane channel and loses its temporary targeting sequence. The core goal of this deck is to show students how mechanical threading and enzymatic cleavage work together simultaneously to process raw biomolecules into functional forms.
Once the ribosome is securely docked at the membrane, the pause in protein synthesis ends, and translation resumes. However, the new amino acids are no longer extruded into the cytoplasm. Instead, the unfolded polypeptide chain is gradually introduced into a specialized transmembrane channel known as the translocon. This structure acts as a protective tunnel, threading the delicate molecule directly into the interior space. The translocon ensures the molecule remains isolated from the chaotic environment of the cytosol while it rapidly elongates.
As the thread passes through the channel, an important modification occurs simultaneously. A specialized enzyme called signal peptidase, located on the inner face of the membrane, recognizes the sequence that initially guided the complex to the organelle. While the machinery of protein synthesis actively pushes new amino acids through the pore, the peptidase snips the signal peptide off the growing chain. This co-translational event highlights how the cell efficiently multi-tasks, modifying the molecule before its construction is even fully complete. The cleaved sequence is subsequently degraded, its job completely finished.
This single enzymatic snip is a transformative event. It converts the initial preprotein into a proprotein, fundamentally altering its structure and destiny. Without the hydrophobic targeting sequence attached, the new proprotein is free to enter the aqueous lumen once translation finishes. This critical conversion step prepares the molecule for the extensive post-translational folding and complex modifications that characterize the final stages of protein synthesis. Failing to cleave this sequence would anchor the molecule improperly, leading to severe cellular blockages and illustrating why every enzymatic step is vital for survival.
Slide 6: Anchoring Protein Synthesis: Integral Membrane Protein Formation
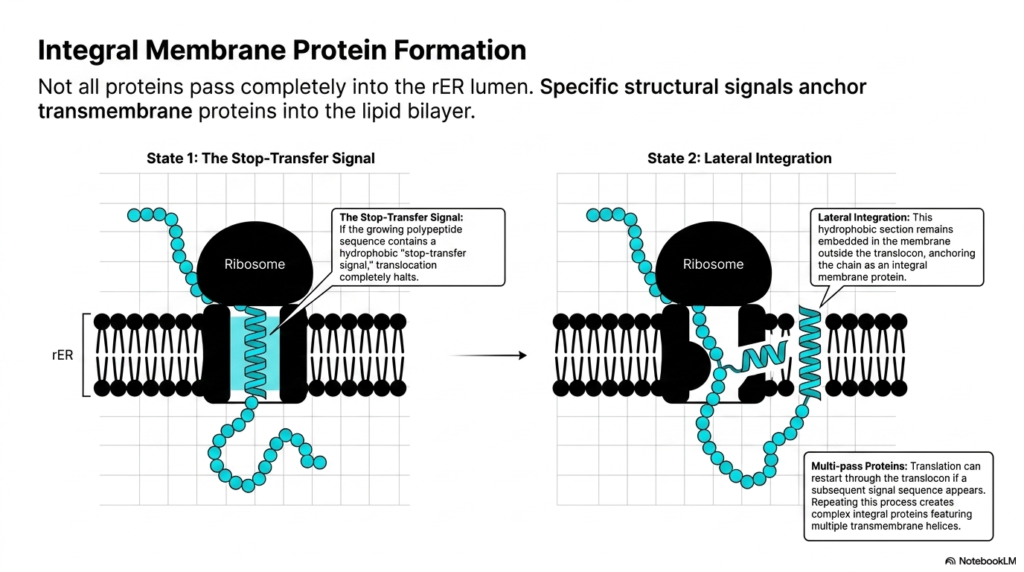
How does a cell build the doors and windows of its own walls? It embeds specialized structures directly into its lipid boundaries. This slide explains how certain molecules become permanently anchored within the membrane rather than passing entirely into the interior space. The objective here is to teach students the structural signals that pause membrane transport, creating the vital receptors and channels cells need to interact with their environment.
Not all products of protein synthesis pass completely through the translocon channel into the luminal space. Many essential cellular components, such as ion channels and hormone receptors, must remain permanently embedded within the lipid bilayer. The formation of these integral membrane proteins relies on specific structural cues built directly into the amino acid sequence. The most critical of these cues is known as the stop-transfer signal. As the ribosome continues its work, this specific sequence dictates the molecule’s final destination.
State that one of these processes begins when the growing polypeptide sequence reveals a dense, hydrophobic stop-transfer signal. When this specific hydrophobic stretch enters the translocon, translocation completely halts. The aqueous pore of the channel rejects the water-repelling segment, causing the forward movement of protein synthesis to pause permanently. The ribosome remains attached to the cytosolic side, continuing to synthesize the remainder of the molecule, but the newly formed chain accumulates in the cytoplasm rather than entering the organelle interior. This mechanism brilliantly allows the cell to position one end of the molecule inside the organelle and the other end outside in the cytoplasm.
State two involves the lateral integration of the molecule into the surrounding lipid environment. The translocon channel physically opens along its side, allowing the hydrophobic section to slip out and remain embedded in the membrane core. This anchors the chain, officially creating an integral membrane protein. For highly complex structures, protein synthesis can actually restart through the translocon if a subsequent signal sequence appears. Repeating this stop-and-start process creates intricate, multi-pass proteins featuring multiple transmembrane helices, a concept crucial for pharmacology students studying drug receptors.
Slide 7: Modifying Protein Synthesis: N-Linked Protein Glycosylation
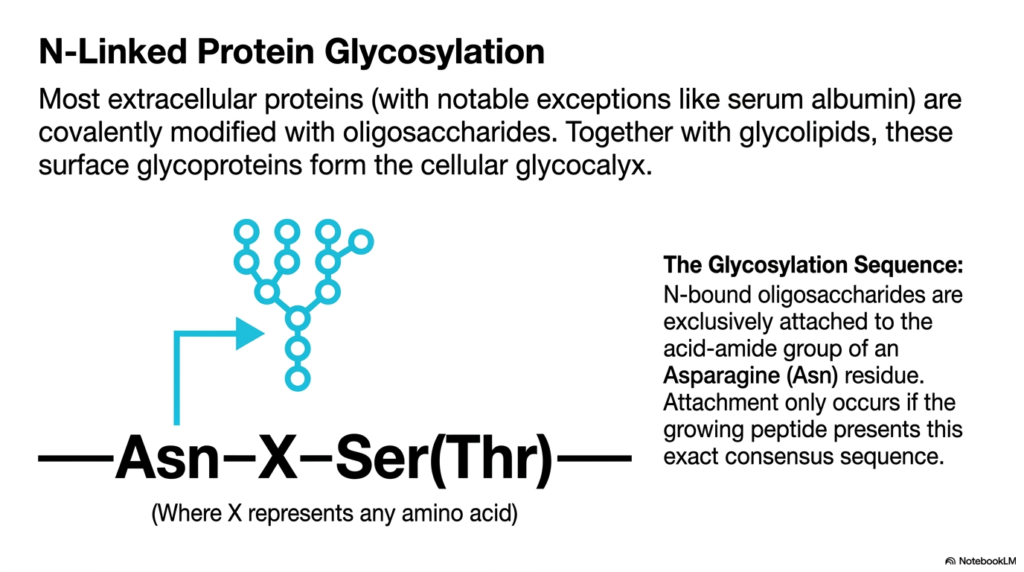
Adding a coat of paint to a newly manufactured car protects it from the elements and defines its final look. Similarly, cells add sugar coats to their molecules to determine their function and stability. This slide introduces N-linked glycosylation, a major modification step. The goal of this presentation is to help students identify the specific consensus sequences that trigger the attachment of carbohydrate branches to new biomolecules.
A significant portion of protein synthesis involves preparing molecules for the harsh environments outside the cell. Most extracellular proteins, with a few notable exceptions such as serum albumin, are covalently modified with complex carbohydrate chains called oligosaccharides. When these surface glycoproteins combine with membrane glycolipids, they form a dense, protective sugar coating around the cell known as the glycocalyx. This glycocalyx is vital for cell-to-cell recognition, immune function, and physical protection, making the study of these modifications essential to medical science. Without these specific sugar modifications, many signaling molecules would rapidly degrade in the bloodstream before reaching their target tissues.
The addition of these sugar chains is not a random event; it requires immense precision. N-bound oligosaccharides are exclusively attached to the acid-amide group of a specific Asparagine residue. However, the presence of Asparagine alone is not enough to trigger the modification. The enzymatic machinery of protein synthesis only acts if the growing peptide presents an exact molecular pattern, known as a consensus sequence. This rigid requirement ensures that only the correct molecules receive these bulky carbohydrate additions.
The required consensus sequence is strictly defined as Asparagine, followed by any amino acid, and ending with either Serine or Threonine. If this exact pattern emerges from the ribosome during protein synthesis, the molecule is immediately targeted for glycosylation. Understanding this sequence allows researchers to predict which parts of a molecule will become glycosylated simply by reading its genetic code. Mutations that alter this sequence can lead to severe congenital disorders, emphasizing its biological importance. For college students, memorizing this consensus sequence is a foundational step in understanding how genetic information translates into complex, three-dimensional cellular structures.
Slide 8: The Scaffold of Protein Synthesis: The Dolichol Diphosphate Anchor
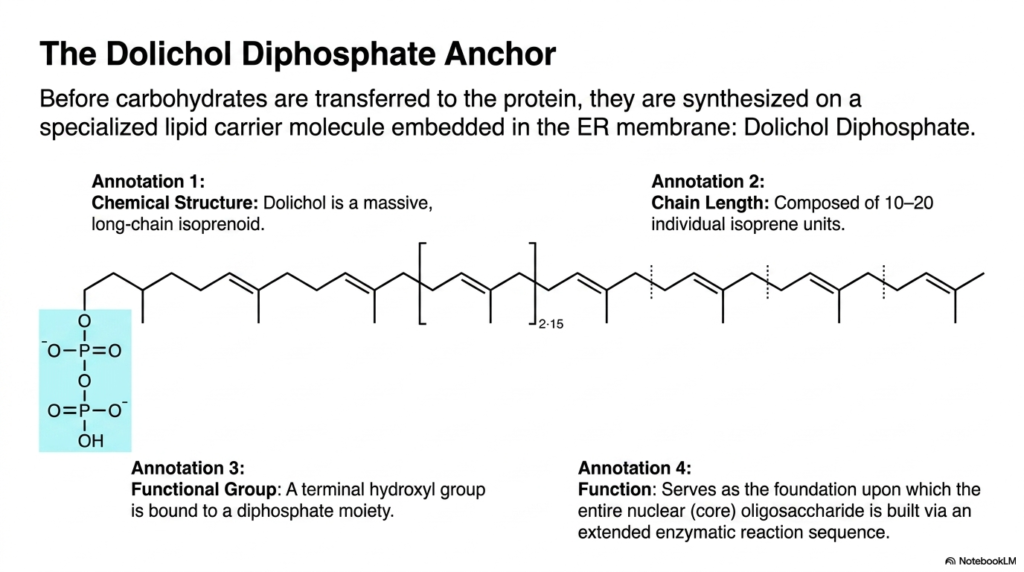
Before constructing a complex roof, builders must first erect a sturdy scaffold. In the cellular factory, massive carbohydrate structures are built on a specialized lipid scaffold before being moved to their final target. This slide examines the chemical structure of Dolichol Diphosphate, the carrier molecule essential for carbohydrate assembly. The purpose of this slide is to introduce students to the unique lipid foundation that supports early molecular maturation.
Carbohydrates are not attached to an emerging polypeptide one sugar at a time. Instead, they are pre-assembled into a massive tree structure before being transferred. This pre-assembly occurs on a specialized lipid carrier molecule embedded directly in the organelle membrane, known as dolichol diphosphate. During the ongoing stages of protein synthesis, this molecule acts as a stable workbench. Understanding its structure is crucial for biochemistry students seeking to visualize how the cell manages the synthesis of bulky, water-soluble carbohydrates in a lipid environment. Without this dedicated workbench, assembling such a massive carbohydrate would be chemically impossible inside the cellular factory.
The chemical anatomy of dolichol is striking. It is a massive, long-chain isoprenoid, specifically composed of 10 to 20 individual isoprene units linked together. This extensive, hydrophobic chain length allows the molecule to anchor itself firmly within the lipid bilayer of the membrane. This deep embedding ensures that the powerful forces involved in protein synthesis and carbohydrate transfer do not tear the carrier from the membrane. It provides the necessary physical stability for the complex enzymatic reactions to follow. The repeating isoprene units create a flexible yet robust tail that integrates seamlessly with the surrounding phospholipids.
The functional end of the dolichol molecule features a terminal hydroxyl group bound to a diphosphate moiety. This highly energetic diphosphate group projects out into the cytoplasm or the lumen, serving as the reactive foundation for the entire process. It is upon this energetic base that the entire nuclear core oligosaccharide is painstakingly built through an extended enzymatic reaction sequence. Grasping the role of this lipid anchor gives students a deep appreciation for the spatial organization and energetic requirements of advanced protein synthesis.
Slide 9: The Blueprint of Protein Synthesis: Anatomy of the Core Oligosaccharide
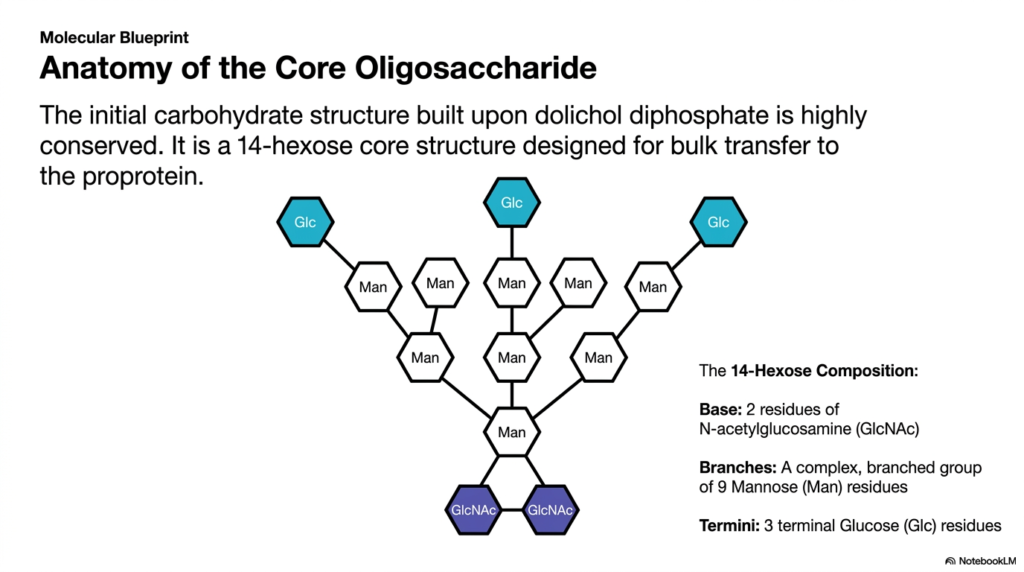
To mass-produce reliable products, factories rely on standardized, interchangeable parts. The cell utilizes a similar strategy by building a universal carbohydrate precursor for its molecules. This slide details the precise molecular blueprint of the core oligosaccharide. The core objective of this deck is to break down this complex sugar tree into its constituent monosaccharides, providing students with a clear structural map of cellular glycosylation.
The initial carbohydrate structure built upon the dolichol diphosphate anchor is remarkably consistent across different life forms. This highly conserved assembly represents a universal starting point for the maturation phase of protein synthesis. Instead of custom-building unique sugars for every new polypeptide, the cell creates a standard 14-hexose core structure. This massive, pre-fabricated sugar tree is specifically designed for bulk transfer to the proprotein. By utilizing a standardized precursor, the cell vastly increases the speed and efficiency of molecular production. This uniformity ensures that quality control mechanisms can easily recognize and process the newly formed chains.
Examining the 14-hexose composition reveals a highly organized and specific architecture. The base of the structure, which attaches directly to the diphosphate anchor, consists of two residues of N-acetylglucosamine. Extending from this rigid base is a complex, extensively branched group composed of nine mannose residues. This branching mannose core provides the bulk of the physical volume and creates numerous attachment points for further modifications. Mastering this specific layout is essential for students tracking the chemical changes that occur during later stages of protein synthesis.
Finally, the outermost tips of the branched structure feature three terminal glucose residues. These terminal sugars are not merely decorative; they serve as critical biological indicators regarding the folding status of the newly formed molecule. The exact arrangement of these fourteen specific sugars represents an incredible feat of enzymatic precision. Understanding this core anatomy provides a vital foundation for medical students studying metabolic diseases, as defects in the assembly of this blueprint during protein synthesis often result in severe, systemic health consequences. Each individual sugar molecule added to the dolichol anchor represents a carefully orchestrated biochemical reaction.
Slide 10: Executing Protein Synthesis: En-Bloc Oligosaccharide Transfer
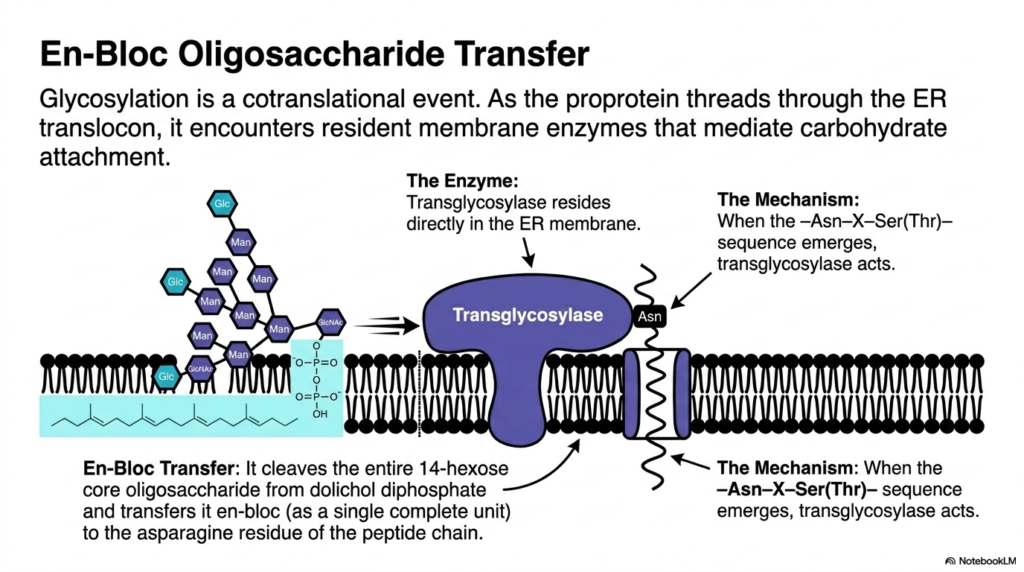
Moving heavy machinery requires specialized cranes capable of lifting entire units at once. In the microscopic cellular world, an enzyme acts as this crane, moving a massive sugar tree in a single, sweeping motion. This slide explores the en-bloc transfer of the core oligosaccharide. The core purpose here is to illustrate the exact enzymatic mechanism by which the complex sugar tree is attached to the newly formed polypeptide chain.
The attachment of the carbohydrate tree is a highly synchronized cotranslational event. This means the modification occurs simultaneously with the ongoing forward movement of protein synthesis. As the newly formed proprotein rapidly threads through the translocon channel, it immediately encounters resident membrane enzymes stationed nearby. These enzymes are primed and ready to mediate the crucial step of carbohydrate attachment. This spatial arrangement ensures that the hydrophobic portions of the chain are shielded and modified before they can improperly fold. The interior of the organelle is a bustling environment where speed and precision are absolutely paramount.
The primary mediator of this event is a massive enzyme known as a transglycosylase, which resides directly within the membrane. Transglycosylase constantly monitors the emerging polypeptide chain. The precise mechanism is triggered only when the specific consensus sequence emerges from the channel’s protective pore. Recognizing this exact pattern, the transglycosylase acts swiftly. This precise timing represents one of the most critical quality control checkpoints in the entire cascade of protein synthesis, ensuring the modification occurs at the correct location. If the enzyme misses this window, the polypeptide may fold in a way that permanently hides the target sequence.
Once triggered, the transglycosylase executes an en-bloc transfer. Instead of moving sugars individually, it cleaves the entire, massive 14-hexose core oligosaccharide away from its dolichol diphosphate lipid anchor. In a single chemical step, it transfers this complete unit directly to the target Asparagine residue of the peptide chain. This bulk transfer method is incredibly efficient, instantly altering the molecule’s physical properties. Understanding this powerful enzymatic action helps biochemistry students appreciate the speed and scale of molecular modifications during protein synthesis.
Slide 11: Refining Protein Synthesis: ER Processing and Oligosaccharide Trimming
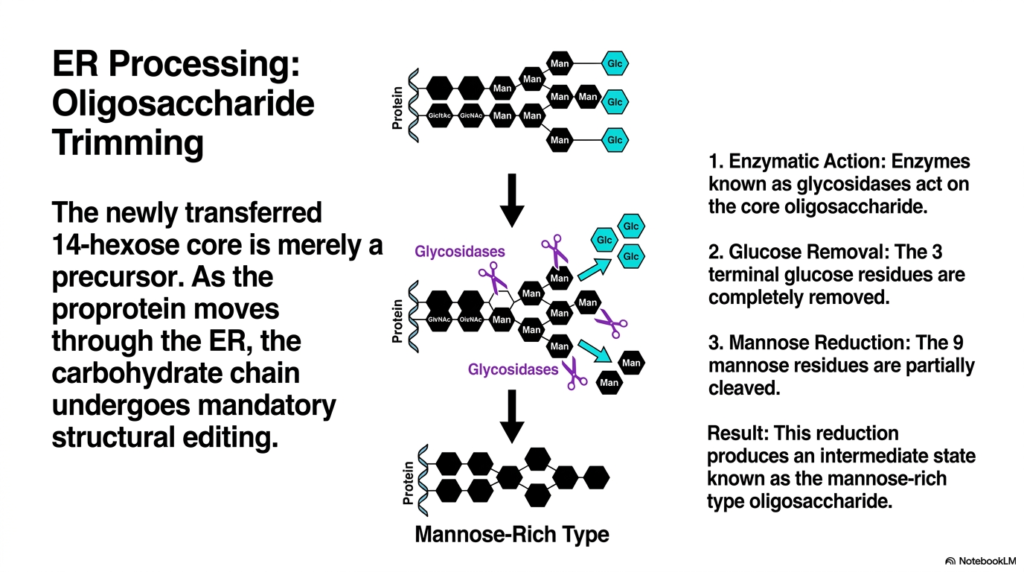
An unpolished sculpture requires careful chipping and sanding to reveal the final masterpiece within. Similarly, the newly attached sugar tree must be immediately trimmed and refined by cellular tools. This slide breaks down the mandatory structural editing that occurs right after carbohydrate attachment. The fundamental goal is to teach students how specific enzymatic actions convert the bulky precursor into a streamlined intermediate.
The en-bloc transfer of the massive 14-hexose core is an impressive feat, but it is only the beginning of molecular maturation. The newly transferred structure is merely a bulky precursor that is not biologically functional in its current state. As the nascent proprotein continues its journey through the organelle during protein synthesis, the carbohydrate chain undergoes mandatory structural editing. This processing stage is vital, as the cell must remove specific sugars to signal that the molecule has folded correctly and is ready for export.
The editing process relies on the swift enzymatic action of specialized proteins known as glycosidases. These enzymes actively seek out and bind to the newly attached core oligosaccharide. The first step in this refinement involves the strict removal of glucose. The glycosidases systematically cleave off the three terminal glucose residues located at the very ends of the branches. This removal acts as a molecular timer in protein synthesis, signaling to the cell’s quality-control sensors that the molecule has successfully attained its proper three-dimensional conformation. If a molecule misfolds, different enzymes will actually reattach a glucose molecule, forcing it back into the folding cycle.
Following the elimination of the terminal glucose units, the enzymatic trimming continues with mannose reduction. The original structure contained nine mannose residues, but the glycosidases partially cleave this branched network. The result of this aggressive enzymatic reduction produces a highly specific intermediate state known as the mannose-rich type oligosaccharide. Students must recognize that this streamlined, mannose-rich structure is the required ticket for the molecule to exit the current compartment and continue the complex pathway of protein synthesis in the Golgi apparatus. This precise trimming process prevents premature transport and ensures that only properly formed structures advance.
Slide 12: Perfecting Protein Synthesis: Golgi Maturation and Complex Glycosylation
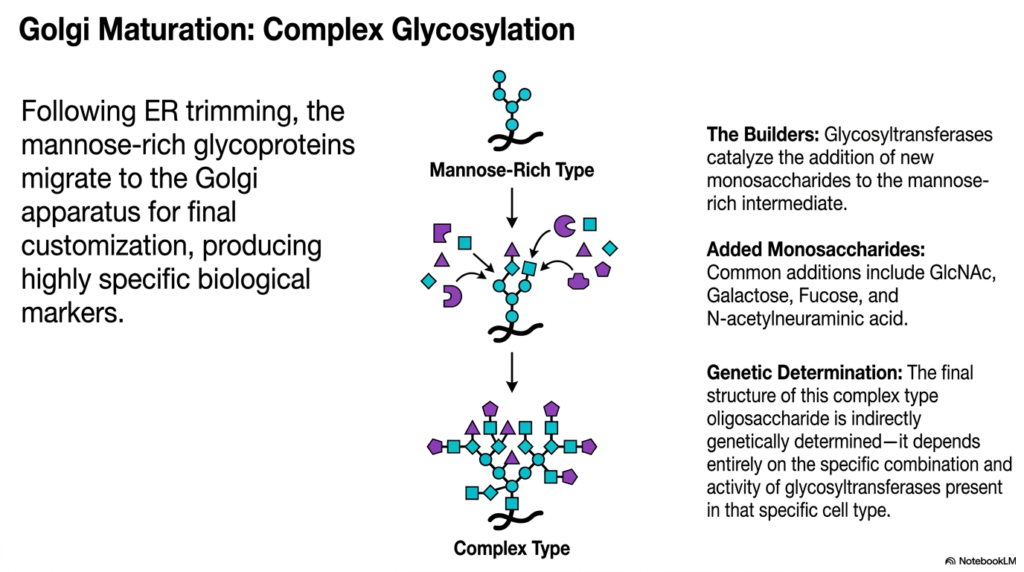
Imagine a custom paint shop adding highly specific decals and finishes to generic vehicles based on their final destination. The Golgi apparatus serves this exact customizing function for cellular molecules. This slide details the transition from a mannose-rich intermediate to a diverse complex type structure. The purpose of this deck is to show students how diverse biological markers are genetically determined and synthesized to dictate final cellular function.
Following extensive trimming in the endoplasmic reticulum, the molecular products of protein synthesis must move to their next processing center. The streamlined, mannose-rich glycoproteins migrate via transport vesicles to the Golgi apparatus. The Golgi serves as the ultimate customization factory, dedicated to producing highly specific biological markers. While early modifications are universally applied to all molecules, this stage of protein synthesis introduces incredible diversity, ensuring that each molecule is specifically tailored to its unique biological role in the body. This is where the generic building blocks are transformed into unique, functional tools.
The builders responsible for this intricate customization are an array of enzymes called glycosyltransferases. These enzymes catalyze the addition of entirely new monosaccharides directly onto the mannose-rich intermediate structure. The additions are incredibly varied and often include specific sugars such as N-acetylglucosamine, galactose, fucose, and N-acetylneuraminic acid. By combining these different monosaccharides in various branching patterns, the cell generates a vast library of complex-type oligosaccharides. This carbohydrate diversity is essential for advanced cellular communication, immune responses, and pathogen recognition.
The final structure of these complex carbohydrate trees is indirectly genetically determined. There is no specific DNA sequence that codes for the exact layout of the sugar branches. Instead, the final product depends entirely on the specific combination, concentration, and activity of glycosyltransferases present within that particular cell type. This means the final stages of protein synthesis look vastly different in a liver cell compared to a neuron. The differential expression of these builder enzymes allows a single organism to produce a staggering array of distinct cellular surfaces. Medical students must grasp this concept to understand blood typing, immunology, and tissue-specific functions.
Slide 13: Decoding Protein Synthesis: The Oligosaccharide Maturation Matrix

Comparing and contrasting concepts side-by-side is one of the most effective ways to master complex scientific material. This slide provides a comprehensive comparative matrix detailing the two major stages of carbohydrate maturation. The core purpose of this slide is to serve as a high-yield study tool, allowing biology students to quickly distinguish between early trimming events and later synthetic additions during the molecular lifecycle.
To truly master the post-translational modifications involved in protein synthesis, students must understand the distinct differences between the two main types of oligosaccharides. The provided matrix clearly outlines the transition from the early mannose-rich type to the final complex type. The location of formation is the first major distinction. Mannose-rich structures are generated exclusively in the endoplasmic reticulum and serve as a universal early-stage milestone. In contrast, the diverse complex types are entirely constructed later within the compartments of the Golgi apparatus. Recognizing these distinct cellular zones helps researchers pinpoint where metabolic errors might occur in disease states.
The primary enzymes at these two locations have entirely opposite functions. In the first compartment, the dominant enzymes are glycosidases. Their enzymatic action relies entirely on cleavage and trimming, systematically breaking down the bulky precursor. Once the products of protein synthesis reach the Golgi, the enzymatic environment shifts radically. Here, glycosyltransferases take over, focusing on the addition and synthesis of new chemical bonds. This shift from destructive trimming to constructive addition represents a fundamental transition in the biochemical pathway.
The monosaccharide composition and biological diversity also change dramatically. The early intermediate is composed solely of N-acetylglucosamine and mannose, representing a highly conserved structure found universally across different cell types. It serves merely as a precursor. The complex type, however, incorporates a wider array of sugars, including galactose and fucose, creating the final functional glycoprotein that makes up the cellular glycocalyx. Grasping this matrix ensures students can accurately track the progression of protein synthesis from a simple chain to a highly specific, diverse molecular tool. This table condenses chapters of textbook material into an accessible, rapid-review format perfect for board exam preparation.
Slide 14: Culminating Protein Synthesis: The Complete Maturation Pathway
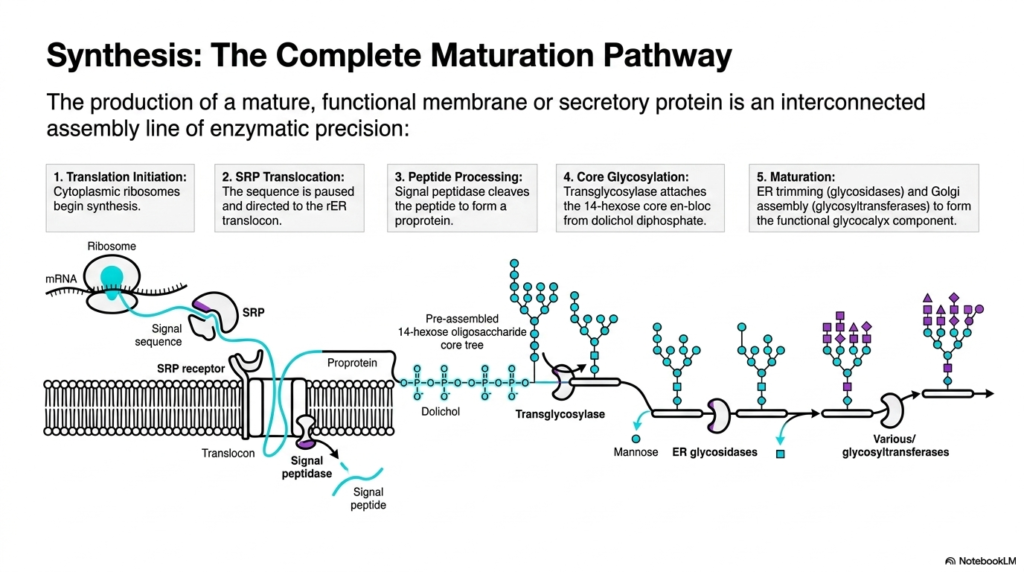
A symphony is only successful when dozens of individual instruments play together in perfect sequential harmony. The production of a functional biomolecule requires a similarly interconnected assembly line of enzymatic precision. This final slide synthesizes the entire journey into a single, comprehensive pathway. The ultimate goal of this presentation is to integrate isolated biochemical concepts into one continuous, logical narrative for advanced students.
Understanding the complete maturation pathway requires viewing protein synthesis as an interconnected assembly line rather than isolated events. The process initiates in the cytoplasm, where ribosomes begin decoding the genetic message. Almost immediately, the signal recognition particle facilitates translocation, pauses the sequence, and safely directs the entire complex to the membrane translocon. This critical hand-off ensures the emerging molecule is shielded from the cytosolic environment. By linking these early steps together, students can visualize the seamless transition from genetic code to physical substance. It demonstrates how mechanical movement and chemical recognition are intricately intertwined.
As the physical threading continues, peptide processing occurs simultaneously. The membrane-bound signal peptidase cleaves the targeting sequence, permanently altering the molecule into a proprotein. Next comes the massive addition of carbohydrates during core glycosylation. The transglycosylase enzyme attaches the pre-assembled 14-hexose core en bloc, utilizing the dolichol diphosphate lipid anchor. These concurrent actions highlight the incredible density and speed of the enzymatic machinery driving protein synthesis. The cell performs major structural edits in milliseconds, long before the entire chain is even fully assembled. This spatial and temporal coordination prevents fatal misfolding errors.
The culmination of this vast assembly line is the final maturation phase. This involves rigorous editing by glycosidases and subsequent construction by glycosyltransferases. The molecule transitions from a generic mannose-rich precursor into a highly specialized component of the functional glycocalyx. Tracking a molecule from translation initiation to its final complex glycosylation state reveals the true elegance of cellular biology. Mastering this entire sequence of protein synthesis empowers medical and biochemistry students to deeply understand the molecular foundations of health, cellular function, and disease pathology. Every single enzymatic checkpoint represents a potential target for modern pharmacological interventions.
Please read our Content Disclaimer Statement.
Check out our social media channels: