76. Endoplasmic Reticulum: Structure, Function, and Mechanics
Imagine a bustling city that never sleeps, constantly manufacturing, modifying, and exporting goods to survive. The human cell operates much like this metropolis, relying on complex internal machinery to sustain life. The following slide deck explores the Endoplasmic Reticulum and Golgi apparatus, the ultimate biochemical factories of the cell. This guide breaks down these fundamental structures for medical and college students, illustrating exactly how cells synthesize proteins, manage lipid metabolism, and maintain internal balance to keep biological systems running smoothly.
1. Introduction to the Endoplasmic Reticulum and Golgi Structure
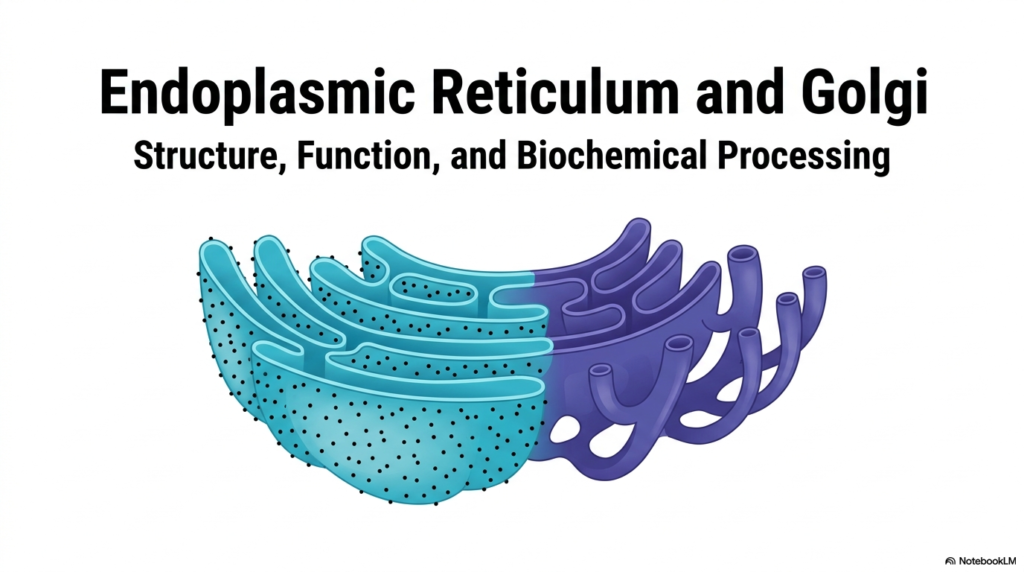
The study of cellular biology begins with understanding the organelles that drive structural and functional organization. The Endoplasmic Reticulum acts as a central pillar in this complex microscopic ecosystem. This initial slide introduces the foundational architecture of the cellular manufacturing plant, highlighting the seamless physical and functional connection between different organelles. It provides a striking visual representation of the interconnected network that underlies all biochemical processing.
Visualizing the Endoplasmic Reticulum alongside the Golgi apparatus reveals a highly organized biochemical assembly line. The intricate folds and tubular extensions demonstrate how physical structure dictates cellular capability. The vibrant contrast in the illustration distinguishes the primary regions of these membrane systems, emphasizing that form and function are deeply intertwined in microscopic spaces. This network is far from static; it represents a dynamic landscape where life-sustaining molecules are constantly formed and refined.
For medical and life science students, grasping the layout of the Endoplasmic Reticulum is crucial for understanding systemic metabolic functions. This system does not just float passively in the cytoplasm; it actively dictates the cell’s biochemical fate. From structural proteins to vital hormones, the sheer volume of biological material that passes through these membranes is staggering. The visual overview sets the stage for a deeper exploration of specialized compartments.
The upcoming sections will dissect these interconnected regions, revealing how the rough and smooth portions of the Endoplasmic Reticulum divide complex labor. By understanding this broad structural overview, observers can better appreciate the localized biochemical events that maintain cellular homeostasis and drive physiological health.
Ultimately, the overarching theme of this presentation is the integration of cellular processes. The Endoplasmic Reticulum stands at the core of this integration, serving as the bridge between genetic instructions and tangible biochemical products. Recognizing this organelle as a continuous, dynamic network rather than a series of isolated structures transforms the way one comprehends cell biology and human physiology.
2. Exploring the Closed Membrane System of the Endoplasmic Reticulum
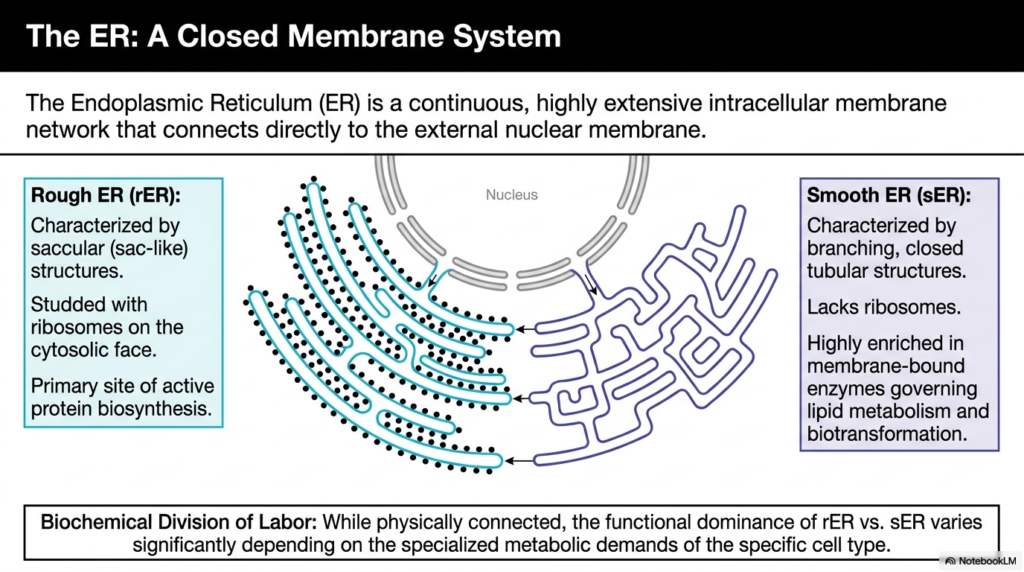
The Endoplasmic Reticulum represents a continuous, highly extensive intracellular membrane network. It physically connects to the external nuclear membrane, allowing genetic information to flow directly into the biochemical production lines. This connection ensures that as mRNA leaves the nucleus, it immediately encounters the machinery necessary for translation. The structural continuity of the Endoplasmic Reticulum underscores its role as the primary interface between the cell’s command center and its manufacturing floors.
This organelle exhibits a remarkable biochemical division of labor, characterized by two distinct structural domains. The rough Endoplasmic Reticulum is identifiable by its saccular, sac-like structures and a cytosolic face heavily studded with ribosomes. These ribosomes are the engines of protein biosynthesis, making this region the primary site for active, targeted protein production. The dense, flattened cisternae provide immense surface area to accommodate thousands of these synthetic complexes simultaneously.
Conversely, the smooth Endoplasmic Reticulum presents a drastically different morphology, characterized by branching, closed tubular structures entirely lacking ribosomes. Instead of producing proteins, this region is highly enriched in membrane-bound enzymes that govern lipid metabolism and cellular biotransformation. The tubular network efficiently houses diverse enzymatic systems required for synthesizing steroid hormones and phospholipids, and for managing vital metabolic pathways across different cell types.
While physically connected, the functional dominance of the rough versus the smooth Endoplasmic Reticulum varies significantly depending on a cell’s specialized metabolic demands. For instance, cells heavily involved in producing protein hormones will exhibit extensive rough endoplasmic reticulum. In contrast, cells responsible for detoxification or lipid synthesis will feature expanded smooth tubular networks. This adaptability makes the entire organelle highly responsive to physiological needs.
Understanding this structural division is foundational for advanced biochemistry. The physical layout of the Endoplasmic Reticulum dictates where and how specific metabolic reactions occur. By compartmentalizing distinct biochemical environments within a single continuous membrane system, the cell achieves remarkable efficiency, preventing cross-contamination between competing metabolic processes while maintaining a streamlined workflow.
3. Protein Biosynthesis at the Rough Endoplasmic Reticulum
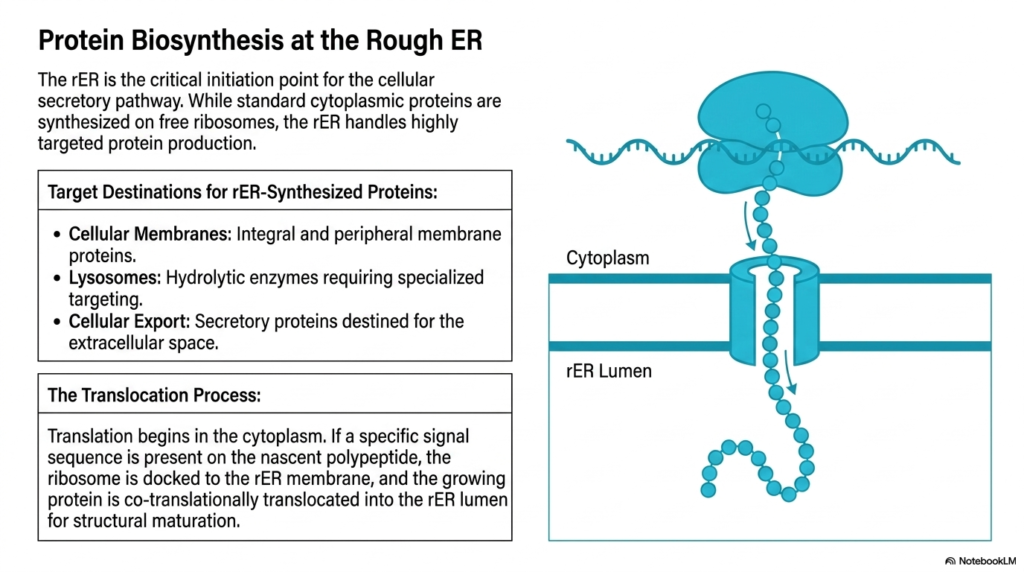
The rough Endoplasmic Reticulum serves as the critical initiation point for the cellular secretory pathway. While standard cytoplasmic proteins are synthesized on free-floating ribosomes, this specific organelle handles highly targeted protein production. It acts as the gatekeeper for macromolecules destined for specialized cellular locations, ensuring that complex biological molecules are built within a controlled environment. The rough Endoplasmic Reticulum isolates these growing polypeptide chains from the general cytoplasm to facilitate precise folding and structural maturation.
Target destinations for proteins synthesized here are highly specific and essential for cellular function. These include integral and peripheral membrane proteins required for structural integrity and cellular signaling. Furthermore, the rough Endoplasmic Reticulum produces hydrolytic enzymes destined for lysosomes, which are critical for degrading cellular waste. Finally, it manufactures secretory proteins slated for export into the extracellular space, such as systemic hormones and digestive enzymes vital for whole-body physiological function.
The translocation process itself is a marvel of biochemical engineering. Translation initially begins in the cytoplasm. However, if a specific signal sequence emerges on the nascent polypeptide chain, the entire ribosome complex is directed and securely docked onto the membrane of the rough Endoplasmic Reticulum. This “address label” mechanism ensures that only the correct proteins enter the secretory pathway, preventing chaotic misrouting of vital metabolic tools.
Once docked, the growing protein is co-translationally translocated directly into the organelle’s protected lumen. The polypeptide chain passes through a specialized channel in the membrane as it is being synthesized. This simultaneous translation and translocation prevent premature folding or aggregation in the cytoplasm. The lumen provides an optimized biochemical environment where the newly minted protein can begin its journey toward functional maturity.
This sophisticated sorting and synthesis system highlights the efficiency of cellular biology. The targeted nature of the rough Endoplasmic Reticulum guarantees that complex structural proteins and highly active enzymes are safely packaged and processed. For medical students, grasping this pathway is fundamental, as defects in ribosomal docking or protein translocation are implicated in numerous metabolic and genetic disorders affecting human health.
4. Post-Translational Maturation inside the Endoplasmic Reticulum
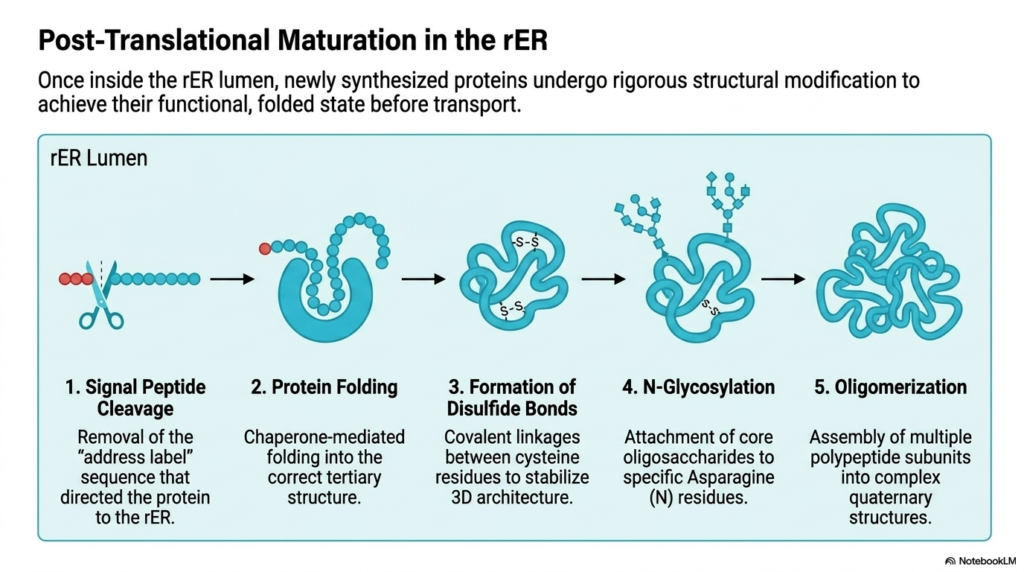
Once a newly synthesized protein enters the lumen of the rough Endoplasmic Reticulum, it must undergo rigorous structural modification. Polypeptide chains are not functional immediately after translation; they require extensive biochemical refinement. This internal compartment provides the precise chemical conditions necessary for proteins to achieve their final, active folded state before transport. The rough Endoplasmic Reticulum acts as a quality control center, ensuring no defective macromolecules proceed further down the secretory pathway.
The first critical step in this maturation process is signal peptide cleavage. The specific “address label” sequence that initially directed the ribosome to the membrane is enzymatically removed by signal peptidases. With this initial tag removed, the protein is permanently committed to the lumen of the rough Endoplasmic Reticulum. Following cleavage, chaperone-mediated folding begins, guiding the linear polypeptide chain into its correct tertiary structural conformation.
Chaperone proteins resident within the Endoplasmic Reticulum prevent improper aggregation and ensure thermodynamic stability. Once folded, enzymes catalyze the formation of disulfide bonds between specific cysteine residues. These covalent linkages act as molecular staples, reinforcing the protein’s three-dimensional architecture. This stabilization is particularly crucial for secretory proteins that will eventually encounter harsh, fluctuating extracellular environments beyond the cell’s protective boundary.
Next, the protein undergoes a process known as N-glycosylation. Core oligosaccharides are enzymatically attached to specific Asparagine (N) residues on the folded protein backbone. This addition of sugar molecules significantly influences protein stability, solubility, and future cellular recognition. Finally, oligomerization occurs, in which multiple modified polypeptide subunits assemble to form complex, multi-part quaternary structures required for advanced biochemical functions.
Only proteins that successfully navigate all these steps are cleared for transport. If a protein fails to fold or glycosylate correctly, the Endoplasmic Reticulum initiates a degradation response to eliminate the faulty product. This rigorous quality control highlights the organelle’s role in preventing cellular toxicity and maintaining the integrity of systemic physiological operations across all biological tissues.
5. Vesicular Transport from the Endoplasmic Reticulum
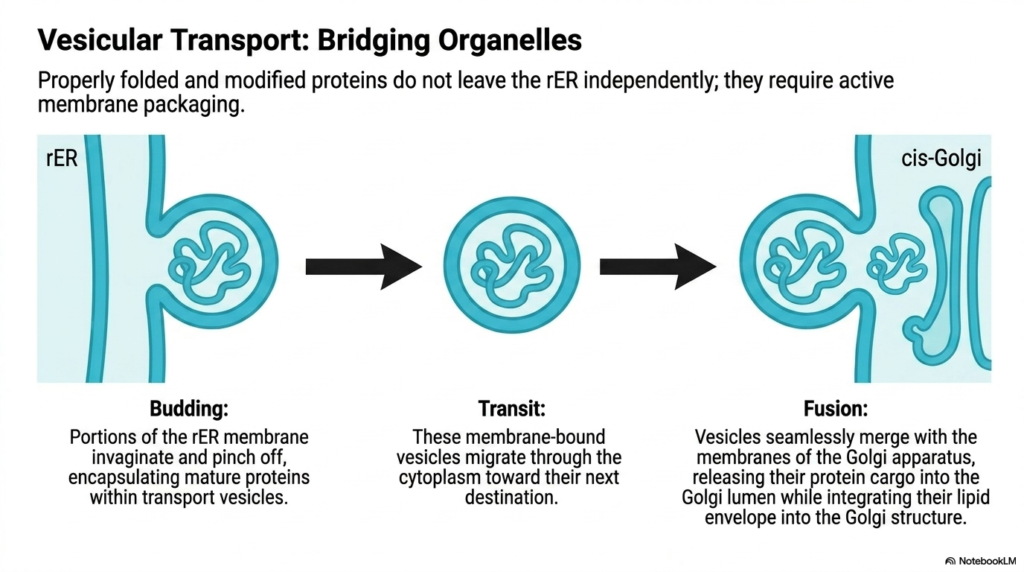
Properly folded and modified proteins cannot leave the Endoplasmic Reticulum independently. They are completely reliant on an active membrane packaging system to safely navigate the treacherous aqueous environment of the cytoplasm. This is where vesicular transport becomes essential, acting as the microscopic cargo system bridging diverse organelles. The rough Endoplasmic Reticulum carefully segregates mature proteins and packages them into protective lipid bubbles designed for precise intracellular delivery.
The transport process begins with a structural event known as budding. Specific portions of the Endoplasmic Reticulum membrane begin to invaginate and pinch off from the main organelle structure. This dynamic membrane remodeling encapsulates the fully mature proteins securely within newly formed transport vesicles. These vesicles represent tiny, mobile fragments of the organelle membrane itself, fully equipped to protect their sensitive biochemical cargo from cytosolic degradation.
Once free, these membrane-bound vesicles enter the transit phase, migrating rapidly through the cytoplasm. Motor proteins drag these vesicles along cytoskeletal tracks, ensuring they travel in a highly directed manner toward their subsequent cellular destination. The vesicles carry not only the internal protein cargo but also the specific lipid composition inherited directly from the Endoplasmic Reticulum, which is crucial for the final step of the journey.
The final stage of this transport mechanism is fusion. Upon reaching the cis-Golgi network, the transport vesicles seamlessly merge with the recipient membranes. The lipid envelope of the vesicle physically integrates into the Golgi structure, while the protein cargo is released directly into the Golgi lumen. This transfer mechanism ensures that sensitive molecules never physically touch the open cytoplasm, maintaining biochemical integrity throughout the logistical process.
Understanding this vesicular transport system is essential for comprehending how cellular compartments communicate and share resources. The Endoplasmic Reticulum initiates this continuous flow of material, driving the dynamic turnover of cellular membranes and the steady supply of functional proteins. Without this precise machinery for budding and fusion, cellular organization would collapse, leading to rapid metabolic failure.
6. Sequential Processing After the Endoplasmic Reticulum
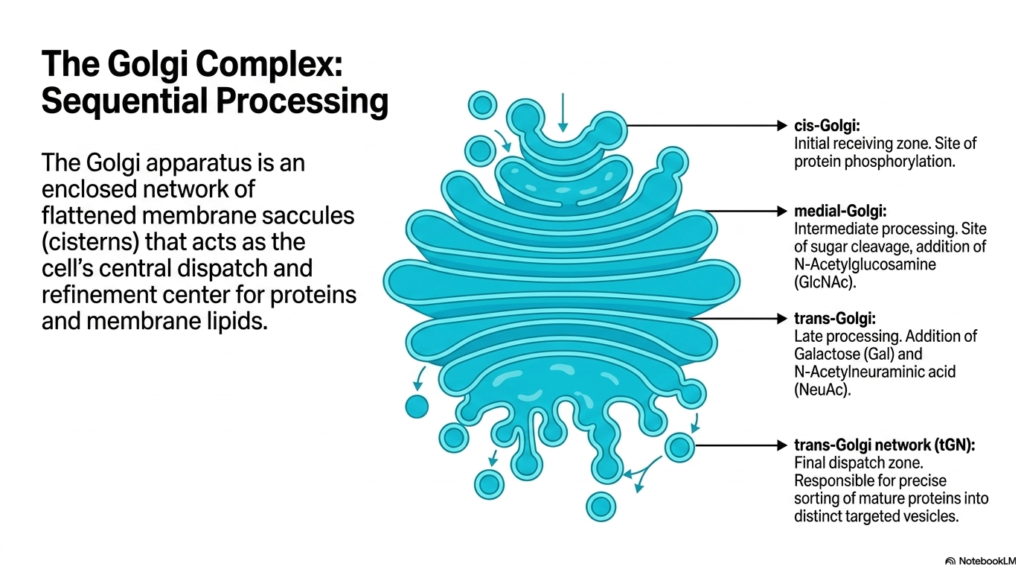
While the Endoplasmic Reticulum acts as the primary synthesizer of proteins, the Golgi complex functions as the ultimate sequential processing and refinement center. The Golgi apparatus is an enclosed network of flattened membrane saccules, or cisterns, serving as the cell’s central dispatch hub. It takes the raw biochemical materials originally manufactured in the Endoplasmic Reticulum and applies specific chemical modifications necessary for their ultimate biological function and cellular targeting.
The processing within the Golgi occurs in a highly organized, step-wise fashion across distinct structural zones. The journey begins at the cis-Golgi, which serves as the initial receiving zone for vesicles arriving directly from the Endoplasmic Reticulum. Within this compartment, proteins undergo initial modifications such as targeted phosphorylation, preparing them for the more complex sugar alterations that occur deeper within the Golgi network.
As proteins move into the medial Golgi, they undergo intermediate biochemical processing. Here, specific enzymes facilitate the cleavage of early sugar modifications and the critical addition of N-Acetylglucosamine. As molecules progress further into the trans-Golgi, late-stage processing occurs, involving the addition of complex sugars such as Galactose and N-Acetylneuraminic acid. This precise, assembly-line modification drastically alters the functional properties of the proteins originally born in the Endoplasmic Reticulum.
Finally, modified molecules reach the trans-Golgi network (tGN), the final dispatch zone. This highly specialized area is responsible for the precise sorting of mature proteins into distinct targeted vesicles. The tGN reads the newly attached biochemical markers to determine the exact cellular destination of every molecule. It serves as the ultimate logistical checkpoint before molecules are permanently dispatched to their functional locations.
The relationship between these two massive organelles is incredibly intimate. The Endoplasmic Reticulum provides the massive bulk of raw polypeptide material, while the Golgi provides the intricate biochemical finesse. Together, they represent a highly integrated, sequential manufacturing process that transforms basic genetic instructions into sophisticated, highly active biological macromolecules required for complex life.
7. Cellular Release Pathways Following Endoplasmic Reticulum Processing
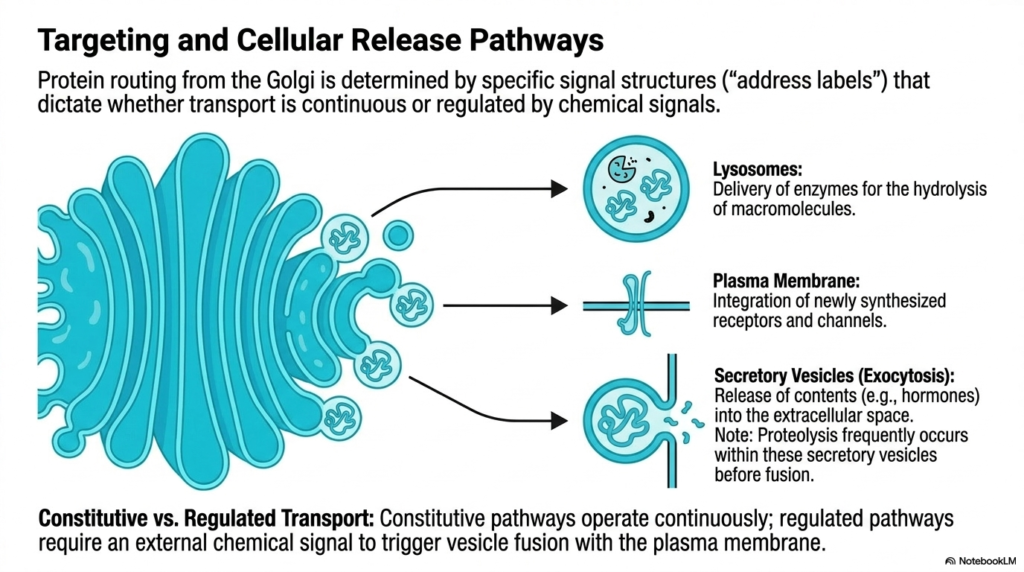
Protein routing from the Golgi complex relies heavily on the specific signal structures initially established during synthesis in the Endoplasmic Reticulum. These intricate biochemical “address labels” dictate exactly where a vesicle must travel and how it should release its cargo. This sorting mechanism ensures that the massive biological output of the Endoplasmic Reticulum is distributed efficiently and correctly, preventing chaotic cellular environments. Transport at this point is strictly divided between continuous flows and highly regulated, chemically triggered pathways.
One major destination for these targeted vesicles is the lysosome. Vesicles routed here carry specialized hydrolytic enzymes designed to degrade macromolecules and cellular waste. Another crucial pathway directs vesicles to the plasma membrane. This route is essential for integrating newly synthesized receptors and ion channels directly into the cell’s outer boundary. By continually supplying these structural components, the Endoplasmic Reticulum indirectly controls cellular communication and environmental sensing.
A third distinct pathway involves secretory vesicles destined for exocytosis. These vesicles carry highly potent contents, such as systemic hormones or digestive enzymes, ready for release into the extracellular space. Interestingly, late-stage proteolysis frequently occurs within these specific secretory vesicles before they ever fuse with the cell membrane. This final cleavage event activates the proteins precisely before they leave the cell, preventing accidental activation during transit.
The cell utilizes two overarching transport strategies to manage these pathways: constitutive and regulated transport. Constitutive pathways operate continuously, providing a steady, uninterrupted flow of newly generated membrane components and extracellular matrix proteins. This baseline transport ensures the constant physical maintenance of the cell membrane and surrounding environment, regardless of external stimuli.
In contrast, regulated pathways stockpile functional molecules and require an external chemical signal to trigger fusion of vesicles with the plasma membrane. For example, insulin produced by the Endoplasmic Reticulum is stored until an influx of glucose signals its rapid, explosive release. This regulated delivery allows the cell to respond dynamically to massive physiological shifts, showcasing the sophisticated logistics of cellular biology.
8. Lipid Metabolism within the Smooth Endoplasmic Reticulum
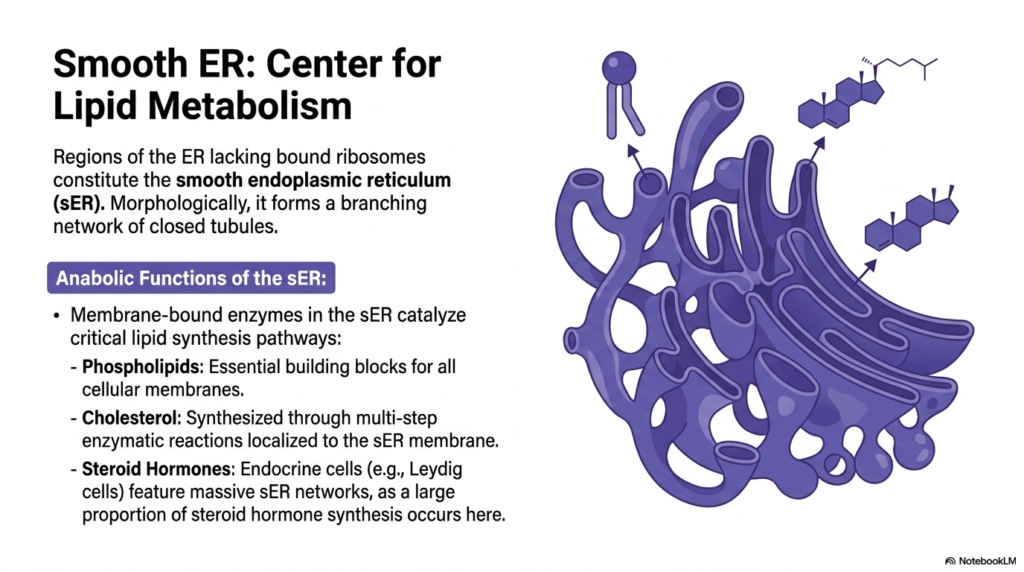
Regions of the organelle lacking bound ribosomes constitute the smooth Endoplasmic Reticulum, an area structurally and functionally distinct from its rough counterpart. Morphologically, this region drops the flattened sac-like appearance in favor of a sprawling, branching network of closed tubules. This unique physical architecture provides massive internal surface area specifically dedicated to housing specialized membrane-bound enzymes. The smooth Endoplasmic Reticulum operates as the indisputable cellular center for complex lipid metabolism and structural fat synthesis.
The anabolic functions localized here are utterly vital for the survival of the cell. Membrane-bound enzymes catalyze critical lipid synthesis pathways, primarily generating phospholipids. These phospholipids are the foundational, essential building blocks required for the creation and repair of all cellular membranes, including the plasma membrane and those of other organelles. By continuously synthesizing these molecules, the smooth Endoplasmic Reticulum ensures the cell’s physical integrity and continued growth.
Beyond basic structural lipids, this compartment also commands the synthesis of complex sterols. Cholesterol is produced here through a highly demanding, multi-step series of enzymatic reactions localized directly to the tubular membranes. This internally synthesized cholesterol is vital for regulating membrane fluidity and serves as the precursor for a massive array of essential biological signaling molecules. The biochemical environment of the smooth Endoplasmic Reticulum enables this energy-intensive synthesis.
Furthermore, this lipid-manufacturing capability is markedly amplified in specialized tissues. Endocrine cells, such as Leydig cells in the testes, feature massively expanded smooth Endoplasmic Reticulum networks. This cellular adaptation exists because a huge proportion of steroid hormone synthesis occurs exclusively within these tubules. These steroid hormones are then released systemically to govern vast physiological networks across the human body.
Understanding this structural specialization is a core concept in medical biochemistry. The complete absence of ribosomes forces this portion of the Endoplasmic Reticulum to abandon protein creation entirely, dedicating all physical and chemical resources to lipid anabolism. This clear biochemical division of labor perfectly illustrates how cellular anatomy dictates physiological capabilities at the microscopic scale.
9. Detoxification Pathways in the Smooth Endoplasmic Reticulum
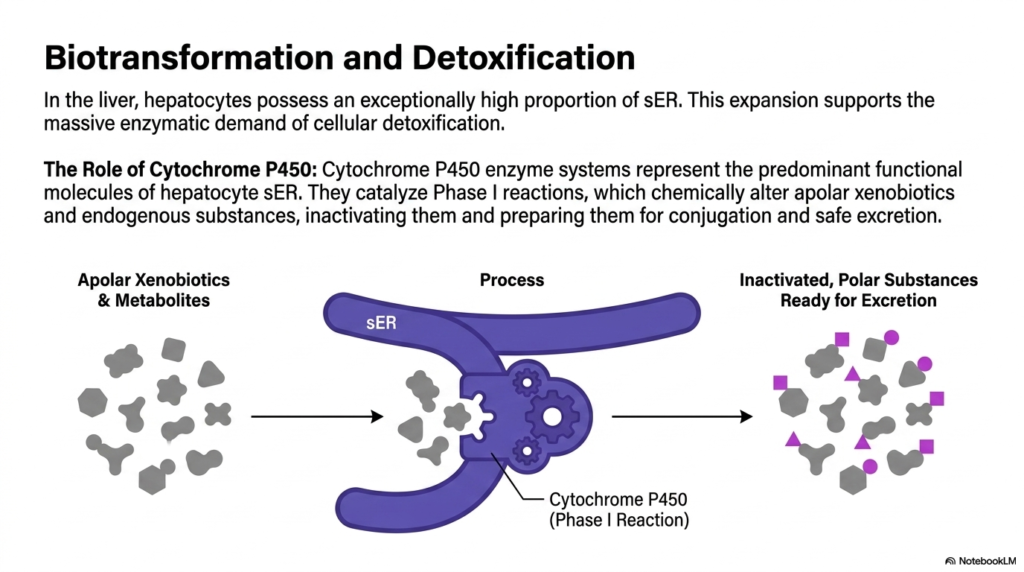
In vital metabolic organs like the liver, hepatocytes possess an exceptionally high proportion of smooth Endoplasmic Reticulum. This massive physical expansion is not for structural support but to sustain the overwhelming enzymatic demands of systemic cellular detoxification. The human body is constantly bombarded by internal metabolic waste and external pharmaceutical agents. The smooth Endoplasmic Reticulum provides the extensive membrane surface area required to house the chemical machinery that neutralizes these potentially lethal toxic compounds.
The absolute biochemical core of this detoxification process relies on the Cytochrome P450 enzyme systems. These proteins represent the predominant functional molecules embedded within the membranes of the hepatocyte smooth Endoplasmic Reticulum. They act as microscopic chemical incinerators, tirelessly processing a vast array of apolar xenobiotics, drugs, and dangerous endogenous byproducts. Without the robust presence of these specific enzymes, toxic molecules would rapidly accumulate and destroy delicate cellular architecture.
Cytochrome P450 enzymes primarily catalyze Phase I detoxification reactions. During this critical process, the enzymes chemically alter dangerous, non-polar substances by introducing reactive polar groups. This targeted oxidation, reduction, or hydrolysis functionally inactivates the foreign compound. The smooth Endoplasmic Reticulum provides an isolated, optimized chemical environment where these volatile, highly reactive chemical transformations can occur without damaging surrounding cytosolic proteins.
By chemically altering these apolar xenobiotics, the enzymes prepare the toxic substances for subsequent Phase II conjugation reactions. The primary goal of these modifications within the smooth Endoplasmic Reticulum is to drastically increase the water solubility of the offending molecule. Converting a lipid-soluble toxin into a polar, water-soluble substance guarantees it can be safely and rapidly excreted from the body via urine or bile.
This extraordinary metabolic capability highlights the dynamic nature of cellular organelles. In individuals frequently exposed to certain drugs or toxins, cells actively synthesize more membrane, radically expanding the smooth Endoplasmic Reticulum to increase detoxification capacity. This morphological adaptation demonstrates how responsive biochemical structures are to systemic physiological stress and environmental challenges.
10. Calcium Storage in the Endoplasmic Reticulum
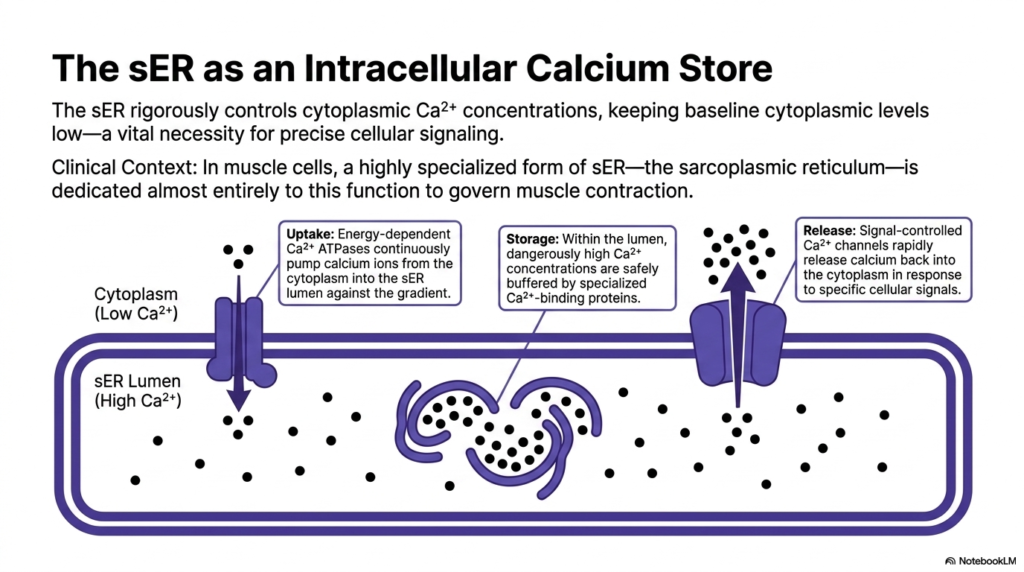
Beyond protein and lipid synthesis, the smooth Endoplasmic Reticulum serves as a critical intracellular calcium store, rigorously controlling cytoplasmic ion concentrations. Maintaining a low baseline level of free calcium in the general cytoplasm is an absolute, vital necessity for precise cellular signaling. If cytosolic calcium levels rise prematurely, it can trigger chaotic, uncontrolled metabolic cascades. The smooth Endoplasmic Reticulum acts as a metabolic vacuum, aggressively sequestering these powerful signaling ions to maintain cellular equilibrium.
In the clinical context of muscle tissue, this organelle undergoes an extreme morphological and functional adaptation. Muscle cells feature a highly specialized form of the smooth Endoplasmic Reticulum known as the sarcoplasmic reticulum. This massive tubular network is dedicated almost entirely to the rapid storage and release of calcium to govern coordinated muscle contraction. Without this massive intracellular ion reservoir, voluntary movement and vital cardiac rhythms would be completely impossible.
The management of this ion gradient requires significant cellular energy. Energy-dependent Calcium ATPases are densely packed into the membrane. These powerful biological pumps continuously force calcium ions from the low-concentration cytoplasm directly into the lumen of the Endoplasmic Reticulum, working aggressively against a steep chemical gradient. This constant, energy-intensive uptake ensures that the resting cytoplasm remains highly sensitive to future signaling events.
Once inside the protective lumen, dangerously high calcium concentrations are safely managed and buffered by specialized, high-capacity calcium-binding proteins. These internal proteins allow the Endoplasmic Reticulum to hold massive quantities of the ion without precipitating destructive internal chemical reactions. The storage capacity is immense, keeping the cellular machinery primed and fully loaded for immediate, explosive signaling when the nervous system physiologically requires it.
When a specific cellular signal arrives, the organelle executes the final phase: rapid release. Signal-controlled calcium channels embedded in the membrane snap open, allowing the stored ions to rapidly flood back into the cytoplasm. This immediate surge triggers downstream physiological responses, from muscle contraction to neurotransmitter release, proving that the Endoplasmic Reticulum is a dynamic control center for cellular communication.
11. Organelle Synthesis Comparison of the Endoplasmic Reticulum
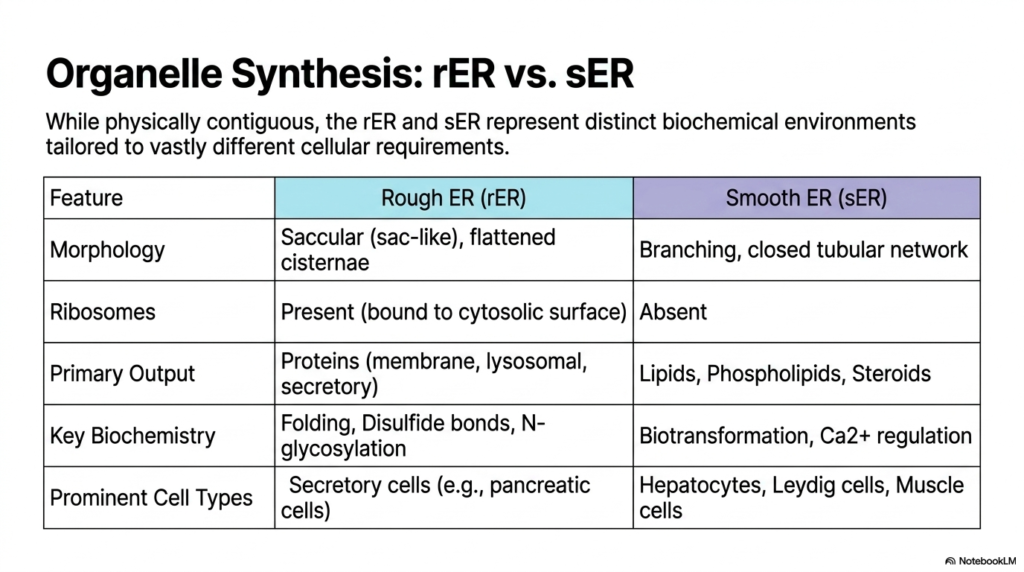
While physically contiguous, the rough and smooth Endoplasmic Reticulum represent radically distinct biochemical environments perfectly tailored to vastly different cellular requirements. Despite sharing a continuous internal lumen and lipid membrane, their functional capabilities are entirely divergent. This comparative slide synthesizes the extreme structural and metabolic differences that allow a single massive organelle to manage nearly all the complex anabolic processes required to sustain life.
The morphological differences are striking and directly facilitate their unique biochemical roles. The rough Endoplasmic Reticulum utilizes saccular, flattened cisternae to maximize surface area for massive arrays of ribosomes bound to its cytosolic surface. In sharp contrast, the smooth Endoplasmic Reticulum completely lacks these ribosomes, organizing itself instead as a highly branching, closed tubular network. This specific tubular architecture provides optimal physical support for large numbers of specialized membrane-bound enzymes and ion channels.
Their primary biological outputs reflect this stark division of labor. The rough region exclusively produces targeted proteins, specifically those destined for cellular membranes, lysosomes, and widespread extracellular secretion. Conversely, the smooth Endoplasmic Reticulum output focuses entirely on lipid synthesis, generating vital phospholipids, cholesterol, and complex steroid hormones. By separating these fundamentally different chemical syntheses into distinct structural domains, the cell prevents chaotic cross-reactions.
Key biochemical processes are heavily compartmentalized between the two regions. The rough Endoplasmic Reticulum is the exclusive site for sophisticated protein folding, the formation of vital disulfide bonds, and early N-glycosylation events. The smooth region commands completely different pathways, driving intense cellular biotransformation, drug detoxification, and rigorous intracellular calcium regulation. Each environment possesses a highly specialized enzymatic toolkit perfectly matched to its specific structural domain.
Finally, the prominence of each region varies drastically depending on the cell type’s overarching function. Secretory cells, like those in the pancreas producing massive quantities of digestive enzymes, are packed with dense rough Endoplasmic Reticulum networks. Meanwhile, hepatocytes managing drug detoxification, Leydig cells synthesizing testosterone, and muscle cells regulating contractile calcium are overwhelmingly dominated by expanded smooth organelle structures.
12. Endoplasmic Reticulum in the Integrated Membrane System
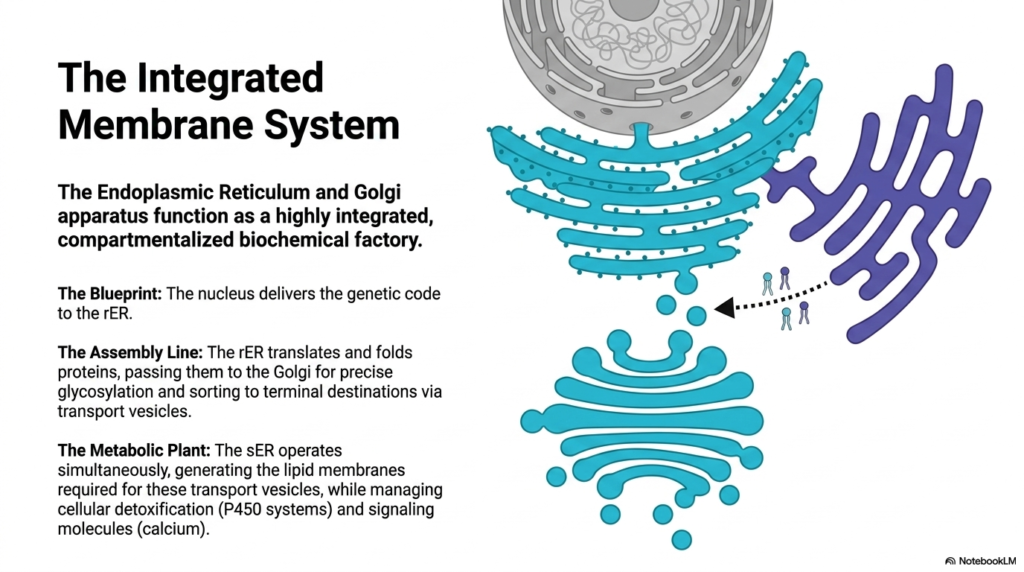
The Endoplasmic Reticulum and Golgi apparatus do not operate in isolation; they function as a highly integrated, compartmentalized biochemical factory. This final slide summarizes the absolute synchronicity required to maintain cellular life, illustrating how distinct microscopic structures collaborate to form a cohesive metabolic pipeline. The entire internal membrane system operates like a masterfully engineered production plant, processing raw genetic data into tangible, highly complex biological molecules essential for physiological survival.
The entire process begins with a precise biological blueprint. The cell’s nucleus stores and protects the genetic code, transcribing it into mobile messenger RNA. This vital code is delivered directly to the surface of the rough Endoplasmic Reticulum. Because the organelle membrane is physically connected to the nuclear envelope, the transfer of genetic information is immediate and highly protected, initiating the massive cellular manufacturing process without delay.
Once the blueprint arrives, the assembly line springs into action. The rough Endoplasmic Reticulum translates the genetic data into raw polypeptide chains, painstakingly folding and stabilizing these new proteins in its protected lumen. It then physically packages these molecules into vesicles and passes them to the Golgi apparatus. The Golgi completes the manufacturing process, executing precise chemical glycosylation and finally sorting the proteins to their terminal biological destinations.
Simultaneously, the metabolic plant operates in the background to sustain this massive logistical effort. The smooth Endoplasmic Reticulum constantly generates new lipid membranes, which are fundamental to creating these countless transport vesicles. Without this continuous lipid production, the entire protein assembly line would quickly grind to a devastating halt, starved of the physical packaging material necessary for intracellular transport.
Furthermore, while supporting this manufacturing pipeline, the smooth Endoplasmic Reticulum concurrently manages cellular detoxification and regulates vital signaling molecules such as calcium. This incredible multitasking demonstrates the profound biological efficiency of the integrated membrane system. Together, these interconnected organelles ensure that the cell can dynamically build, protect, and regulate the complex biochemical components required for human life.
Please read our Content Disclaimer Statement.
Check out our social media channels: