75. Membrane Receptors: Principles of Signal Transduction
Imagine a fortress surrounded by high walls, unable to see the outside world yet perfectly coordinating its defenses against unseen threats. The human cell operates similarly, relying entirely on sophisticated molecular sentinels to perceive and react to its environment. This presentation explores the complex architectures and mechanisms of these sentinels. The core purpose of this slide deck is to break down the biochemical pathways that govern signal transduction, equipping future scientists and medical professionals with a comprehensive understanding of how cells communicate, survive, and thrive.
Slide 1: Introduction to Membrane Receptors and Signal Transduction
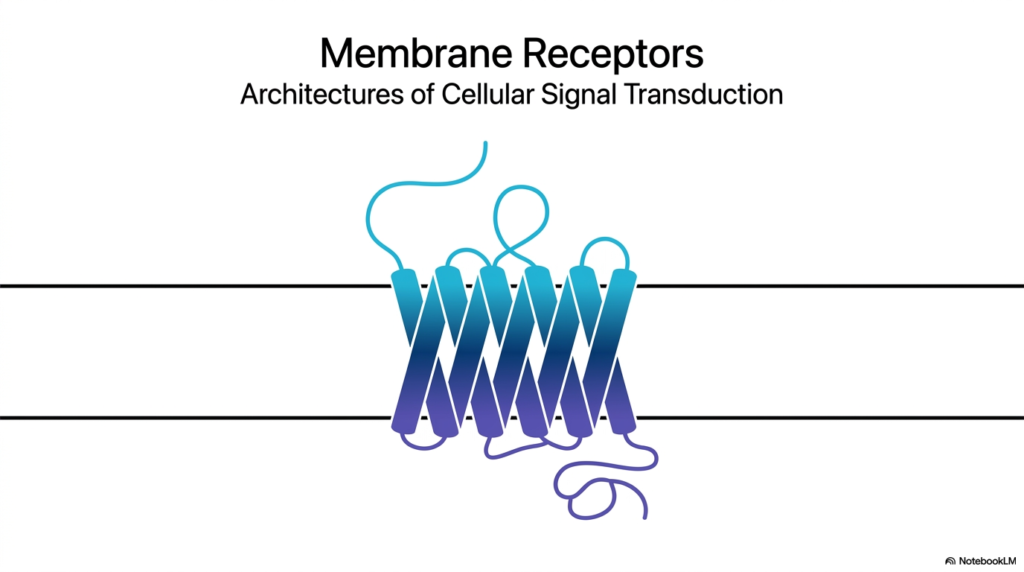
The introductory visual slide presents a striking illustration of a seven-helix transmembrane protein, a classic representation of Membrane Receptors. These complex molecular machines are the fundamental gateway for cellular communication, bridging the external environment and the cell’s internal biochemical machinery. Without Membrane Receptors, cells would exist in total isolation, completely oblivious to hormones, neurotransmitters, and critical environmental changes. This foundational image captures the essence of cellular signal transduction, depicting a physical conduit through which invisible messages pass. Membrane Receptors establish the architectural blueprint for how biological information transforms from a physical or chemical stimulus into an actionable cellular command.
In the study of biochemistry and physiology, understanding Membrane Receptors is absolutely critical. They act as translators, converting an extracellular language into an intracellular one. The protein structure shown on the slide, winding back and forth across the lipid bilayer, highlights a remarkable evolutionary design. The specific folding of the polypeptide chain creates distinct domains that are uniquely capable of capturing signals outside the cell and transmitting that information across the impermeable lipid barrier. This structural complexity is what allows cells to execute highly coordinated functions across entire tissues and organs, maintaining the delicate balance of homeostasis required for human health.
This introductory slide sets the stage for a deep exploration into the diverse forms and functions of Membrane Receptors. The subsequent slides will dissect the specific mechanisms by which different receptor classes operate, ranging from simple single-helix structures to intricate multi-protein complexes. Students of biology and medicine must master the architectures of these proteins, as their dysfunction is at the root of countless human diseases, from diabetes to cancer. By unlocking the structural principles of these biological sentinels, one gains profound insight into the targeted therapies and pharmacological interventions that define modern biochemical medicine.
Slide 2: Anatomy of Cellular Communication via Membrane Receptors
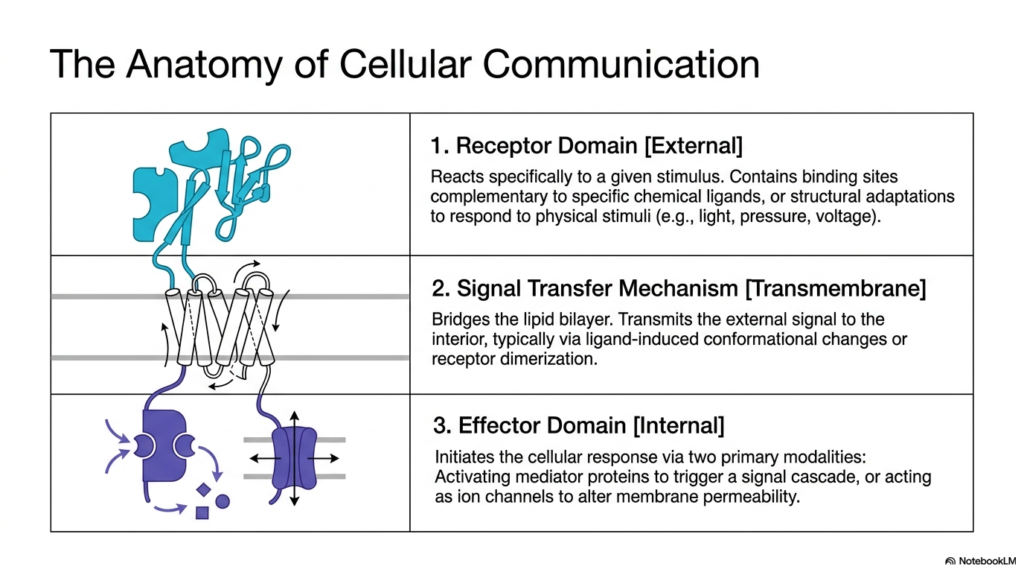
To truly understand how cellular signals propagate, one must first dissect the fundamental anatomical regions of Membrane Receptors. The second slide outlines a tripartite structural model that defines almost all cellular communication pathways. At the outermost boundary lies the external receptor domain. This specialized region is highly selective, featuring binding pockets that perfectly complement specific chemical ligands or possess structural adaptations to capture physical stimuli such as light or pressure. It acts as the initial sensor. When this domain encounters its specific trigger, the communication sequence begins, demonstrating the exquisite specificity that Membrane Receptors possess in distinguishing relevant signals from surrounding noise.
Beneath the external sensor lies the signal transfer mechanism, which represents the transmembrane segment of Membrane Receptors. This domain physically bridges the cell’s impermeable lipid bilayer. Because the chemical messenger often cannot cross the cell membrane, the receptor must transmit the signal mechanically. This transfer typically occurs via profound ligand-induced conformational changes or via receptor dimerization. The protein’s physical shape shifts, carrying the message across the boundary without allowing the extracellular molecule itself to enter. This elegant mechanical translation ensures that the internal cellular environment remains protected while still receiving vital external instructions.
The final crucial component is the internal effector domain, which dictates the ultimate cellular response. Once the transmembrane segment shifts, it activates this intracellular region. The effector domain generally operates via two primary modalities to execute its function. It may activate specific mediator proteins that trigger an extensive internal signaling cascade, or it may act directly as an ion channel to rapidly alter the electrical permeability of the cell membrane. This functional breakdown reveals why Membrane Receptors are the indispensable engines of cellular response, driving everything from metabolic adjustments to rapid muscle contractions.
Slide 3: Stimulus Typology and the Triggers of Membrane Receptors
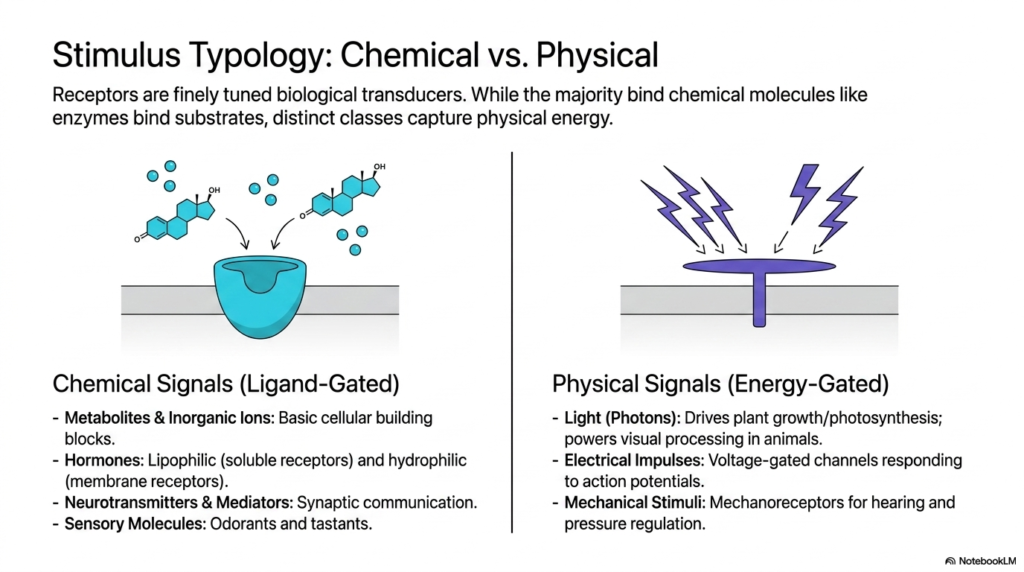
The biological world is rich with diverse stimuli, and Membrane Receptors function as finely tuned biological transducers to interpret this vast array of information. The third slide distinguishes between two primary stimulus categories: chemical and physical. On one side, chemical signals activate ligand-gated receptors. Much like enzymes interact with highly specific substrates, these Membrane Receptors bind to discrete chemical molecules. This category encompasses a broad spectrum of vital biological messengers, including basic metabolites, inorganic ions that serve as cellular building blocks, and hormones. It also includes neurotransmitters that enable rapid synaptic communication in the nervous system, as well as sensory molecules such as odorants and tastants.
While chemical messengers dominate much of cellular communication, a distinct and equally fascinating class of Membrane Receptors captures physical energy. These energy-gated receptors do not rely on a free-floating molecule. Instead, they translate fundamental physical forces into biochemical language. The most famous examples include photoreceptors in the eye, which use specific proteins to capture incoming photons of light, driving visual processing in animals and fueling photosynthesis in plants. These functional adaptations highlight the remarkable evolutionary versatility of the receptor architecture. This incredible capability allows organisms to interact seamlessly with their environment, reacting to subtle shifts in energy that dictate behavior and survival.
Furthermore, the realm of physical signals extends to electrical impulses and mechanical forces. Voltage-gated channels are specialized Membrane Receptors that respond directly to electrical action potentials, a mechanism fundamental to cardiac rhythms and neural firing. Mechanoreceptors add another layer of sensory perception, regulating pressure and enabling hearing by translating physical deformation into cellular signals. By categorizing these diverse triggers, the slide emphasizes that regardless of whether a signal is a chemical hormone or a physical photon, a dedicated receptor exists to translate that specific stimulus into actionable biological intelligence.
Slide 4: The 1-Helix Architectures of Specific Membrane Receptors
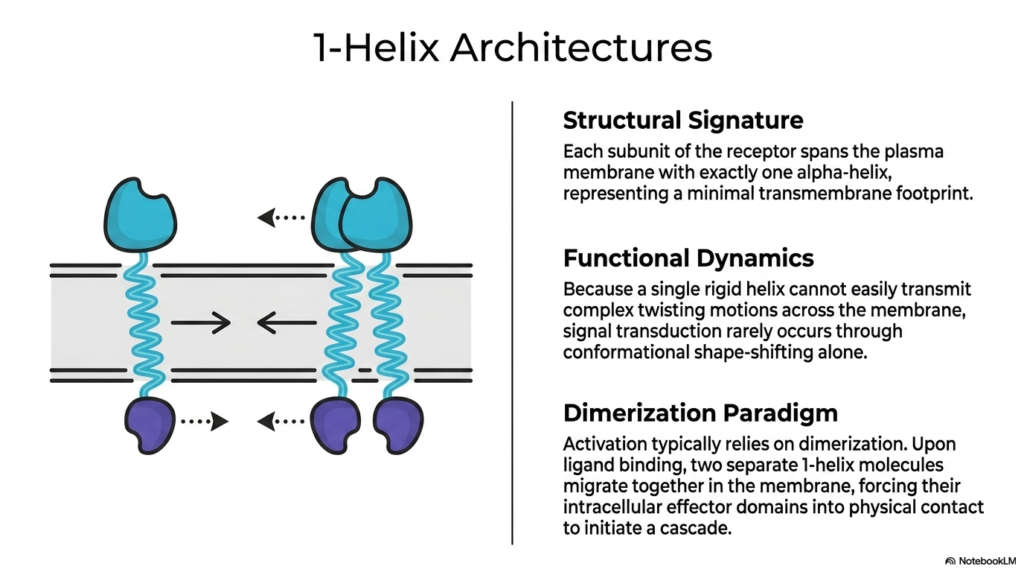
Delving deeper into structural variations, the fourth slide introduces the streamlined design of 1-helix architectures. The defining structural signature of these specific Membrane Receptors is quite literal: each subunit of the receptor protein spans the plasma membrane using exactly one alpha-helix. This represents the absolute minimal transmembrane footprint possible for a functional receptor. Unlike complex, multi-spanning proteins, this simple solitary helix presents a unique biomechanical challenge. The functional dynamics of a single rigid helix dictate that it cannot easily transmit complex twisting or rotational motions across the cellular membrane to convey an internal chemical signal.
Because isolated conformational shape-shifting is highly restricted in such a rigid and simple structure, these single-spanning Membrane Receptors rarely rely on internal structural twisting to communicate. Instead, their activation relies heavily on an elegant mechanism known as the dimerization paradigm. In the absence of a chemical signal, these receptor subunits frequently float independently within the fluid lipid bilayer. However, upon the arrival and successful binding of a highly specific external ligand, a dramatic physical reorganization occurs. The binding event compels two separate 1-helix molecules to migrate toward one another and physically pair up.
This lateral migration and subsequent pairing are the pivotal steps for signal transmission in these Membrane Receptors. By forcing the two subunits together on the cell surface, their respective intracellular effector domains are simultaneously drawn into close physical contact within the cell. This forced intracellular proximity initiates a potent biochemical cascade. It serves as a brilliant evolutionary solution, utilizing lateral movement within the membrane to overcome the rotational limitations of a single transmembrane helix. This specific physical interaction ultimately drives vital physiological processes such as cell growth and metabolic regulation.
For medical and college students, mastering the mechanics of these 1-helix architectures is incredibly important. By grasping how these specific proteins overcome their structural simplicity through dimerization, researchers can better design pharmaceutical drugs that either promote or block these physical pairings, thereby fine-tuning cellular behavior.
Slide 5: The Insulin Receptor as a Model for Membrane Receptors
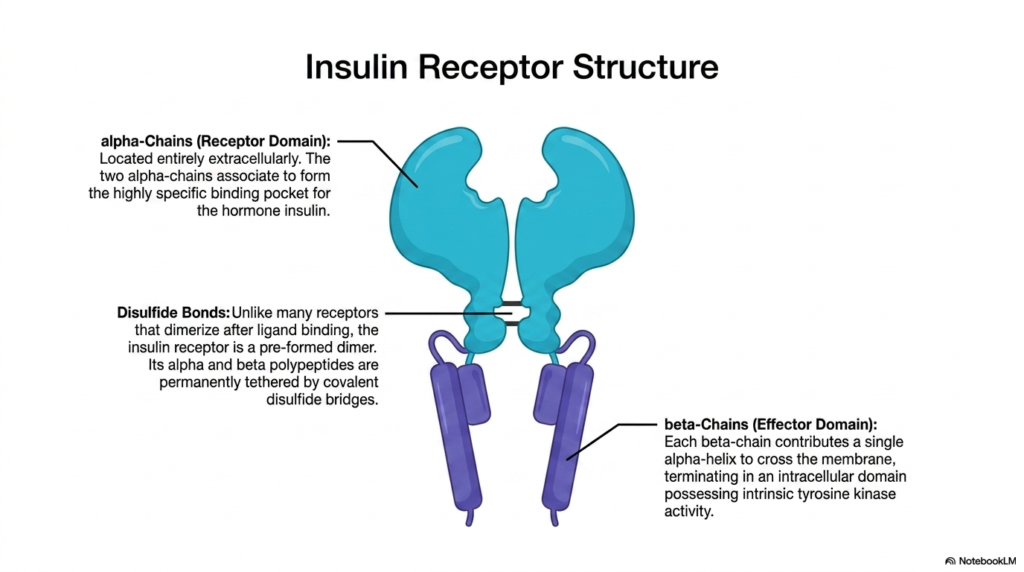
The fifth slide provides a concrete, highly significant biological example of single-helix architectures by detailing the insulin receptor. Among the many diverse Membrane Receptors in the human body, the insulin receptor stands out due to its unique structural adaptations and its profound role in human metabolism. The receptor features prominent alpha-chains that comprise the dedicated receptor domain. Crucially, these alpha-chains are located entirely on the extracellular side of the cell. These two chains associate closely to form a highly specific and complex binding pocket designed exclusively to capture the circulating hormone insulin.
A fascinating structural deviation occurs with the insulin receptor compared to other proteins in its class. Unlike many typical Membrane Receptors that only dimerize after a ligand binds, the insulin receptor exists as a pre-formed dimer. The slide highlights that its distinct polypeptide components are permanently tethered together by strong, covalent disulfide bridges. These disulfide bonds ensure that the structural integrity of the complex remains incredibly stable, primed and ready for immediate action the moment insulin appears in the bloodstream, allowing for rapid metabolic responses.
Anchoring this massive extracellular structure to the cell are the beta-chains, which function as the critical effector domain. Each beta-chain contributes a single alpha-helix to physically cross the lipid bilayer, demonstrating the classic one-helix architecture. However, the most important feature lies at the intracellular termination of these beta-chains. Here, the chains possess specialized domains with intrinsic tyrosine kinase activity. In these specific Membrane Receptors, the enzyme is built directly into the receptor itself. This built-in kinase is the biochemical engine that ultimately drives the cellular response to insulin, thereby regulating blood sugar dynamics.
This comprehensive architectural map of the insulin receptor is a foundational concept in metabolic biochemistry. Understanding the precise relationship between the extracellular binding pocket and the internal kinase domain allows future physicians to grasp exactly how insulin resistance develops and how modern therapeutics might intervene to correct these essential metabolic pathways.
Slide 6: Signal Transduction and Autophosphorylation in Membrane Receptors
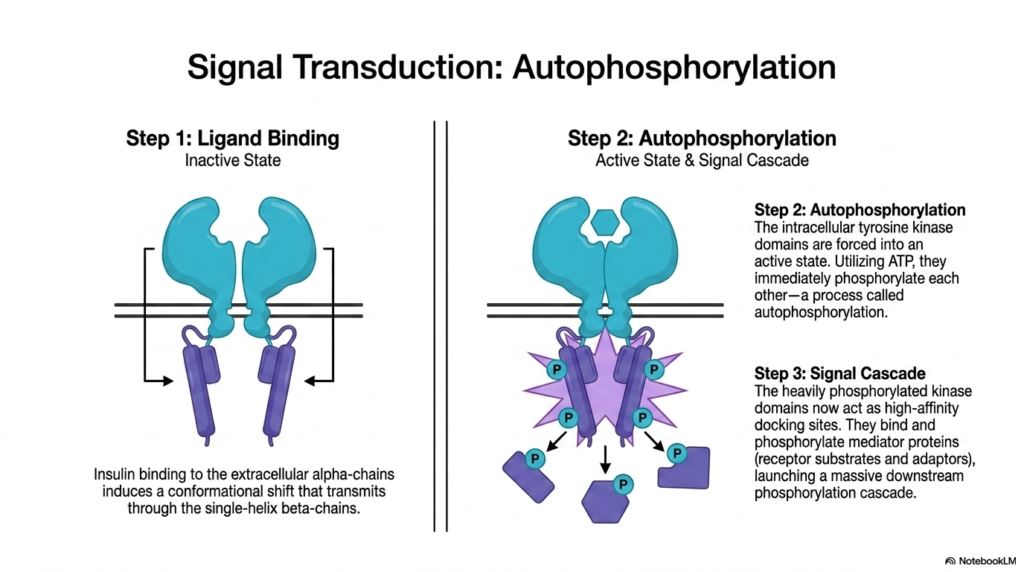
Having established the structural blueprint, the sixth slide illuminates the dynamic functional sequence of the insulin receptor, serving as a prime example of signal transduction. The process begins with Step One: Ligand Binding. In the inactive state, the receptor awaits a signal. When insulin successfully binds to the extracellular alpha-chains, it induces a critical conformational shift. This precise mechanical movement transmits a force directly down through the single-helix beta-chains. This step vividly demonstrates how Membrane Receptors act as mechanical levers, translating an external chemical docking event into a physical movement across the cellular membrane.
This transmembrane shifting forces the receptor into Step Two, a fascinating biochemical event known as autophosphorylation. The physical movement forces the intracellular tyrosine kinase domains of the pre-formed dimer into a highly active state. Using ambient cellular ATP, these kinase domains immediately begin to phosphorylate one another. They attach high-energy phosphate groups to specific tyrosine residues on their partner chain. In the study of Membrane Receptors, autophosphorylation represents the exact moment a signal shifts from a mere physical transmission into a potent biochemical activation, fundamentally changing the chemical properties of the internal receptor domain.
Finally, Step Three details the resulting massive Signal Cascade. The newly and heavily phosphorylated kinase domains undergo a functional transformation, now acting as high-affinity docking sites for various intracellular proteins. These specific Membrane Receptors recruit mediator proteins, which are often receptor substrates and adaptors, binding them tightly and phosphorylating them in turn. This launches an extensive downstream phosphorylation cascade, acting like a biochemical domino effect. A single molecule of insulin bound to the exterior surface rapidly amplifies into thousands of activated internal proteins, swiftly commanding the cell to absorb glucose from the blood.
Grasping the nuance of autophosphorylation and the subsequent cascade is imperative for biochemistry students. It reveals how an incredibly small amount of a hormone can generate a massive, immediate, and highly coordinated cellular response, ensuring metabolic stability across the entire organism in real time.
Slide 7: Exploring the 7-Helix Architectures of Membrane Receptors
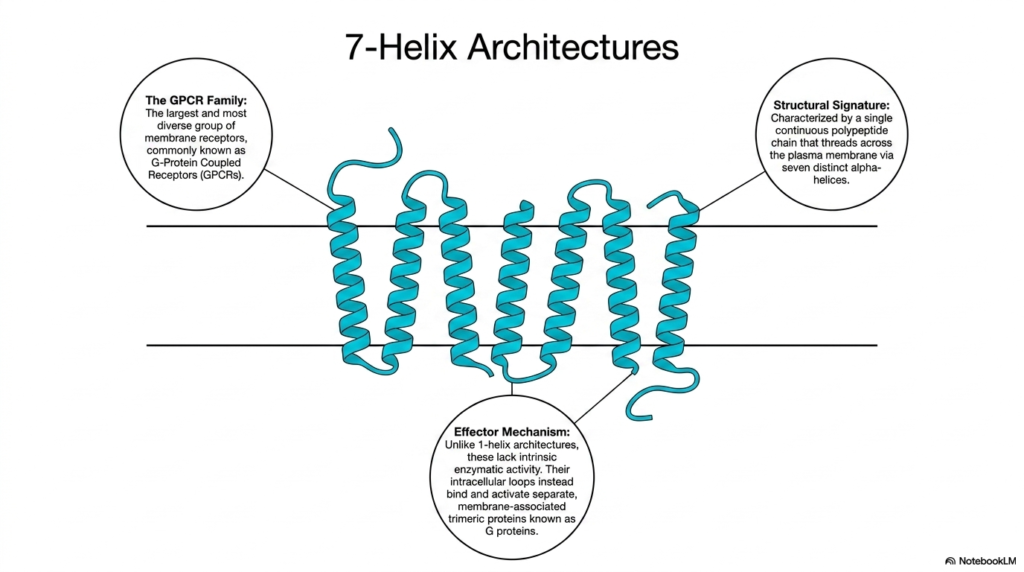
Shifting focus away from the single-helix models, the seventh slide introduces the most diverse and expansive group of cellular sensors: the 7-helix architectures. Biologists and pharmacologists know this massive family as the G protein-coupled receptors, or GPCRs. These specific Membrane Receptors represent the largest class of sensory and communication proteins in the human genome. They are incredibly prolific, responsible for regulating nearly every known physiological process, from heart rate and blood pressure to mood regulation and sensory perception, making them the targets of the vast majority of modern medications.
The structural signature of GPCRs is incredibly distinct and highly conserved across species. Unlike the single-spanning proteins discussed previously, these Membrane Receptors are characterized by a single, continuous polypeptide chain that elegantly threads itself back and forth across the plasma membrane precisely seven times. This creates a complex, serpentine structure consisting of seven distinct transmembrane alpha-helices. This interwoven, barrel-like shape provides immense structural stability and creates numerous internal and external loops. These loops serve as highly customizable interaction sites for capturing a massive array of different ligands and signaling molecules.
The effector mechanism of these 7-helix structures differs fundamentally from that of their 1-helix counterparts. While insulin receptors possess built-in kinase enzymes, GPCR Membrane Receptors lack any intrinsic enzymatic activity of their own. They cannot directly phosphorylate other proteins. Instead, their crucial intracellular loops are designed to physically bind and activate separate, membrane-associated helper molecules known as trimeric G proteins. The receptor acts solely as the activator switch, relying entirely on these associated G proteins to carry the biochemical message further into the cellular interior and initiate the desired metabolic or genetic changes.
This modular design separates the signal detection mechanism from the enzymatic signal execution. This allows cells to mix and match different receptors with various internal G proteins, creating a highly adaptable and complex internal communication network that students must understand to map cellular signaling pathways accurately.
Slide 8: The G Protein Complex and Membrane Receptors in Visual Transduction
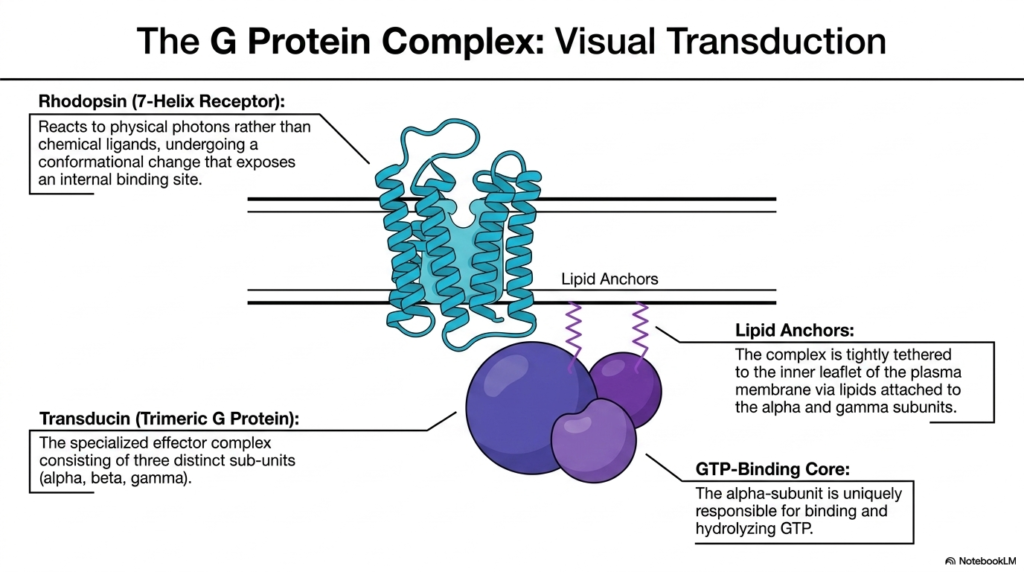
To illustrate the functional reality of 7-helix GPCRs, the eighth slide focuses on visual transduction, examining the magnificent partnership between the receptor Rhodopsin and its dedicated G protein, Transducin. Rhodopsin acts as a highly specialized member of the membrane receptor family, located within the retina. Interestingly, instead of binding a chemical ligand in the extracellular fluid, Rhodopsin is precisely tuned to respond to light photons. When a photon strikes, it forces Rhodopsin to undergo a sudden and profound conformational change, thereby exposing a critical binding site on its intracellular face.
Waiting just inside the membrane is the specialized effector complex known as a Trimeric G Protein, specifically named Transducin in this visual pathway. As the name suggests, this complex consists of three distinct subunits: alpha, beta, and gamma. These proteins do not float freely in the deep cytoplasm. Instead, the entire trimeric complex is tightly tethered to the inner leaflet of the plasma membrane. Lipid anchors, essentially fatty acid chains attached specifically to the alpha and gamma subunits, embed themselves into the lipid bilayer, ensuring the G protein remains perfectly positioned next to the Membrane Receptors.
At the heart of this complex lies the GTP-Binding Core, which is localized entirely within the alpha-subunit. This specific alpha-subunit holds the unique responsibility for binding and eventually hydrolyzing a molecule of Guanosine Triphosphate (GTP). When the activated Rhodopsin physically interacts with the tethered Transducin complex, it targets this alpha-subunit, prompting it to exchange its bound nucleotides. This intricate molecular dance demonstrates how Membrane Receptors physically manipulate secondary complexes to translate an ephemeral flash of light into a permanent, chemical signal that the brain will eventually recognize as vision.
The study of this complex reveals the elegant spatial organization required for cellular signaling. By utilizing lipid anchors to keep all the necessary biochemical components localized in the same two-dimensional plane of the membrane, the cell drastically increases the speed and efficiency of the visual response.
Slide 9: The Amplification Cycle of G-Protein Coupled Membrane Receptors
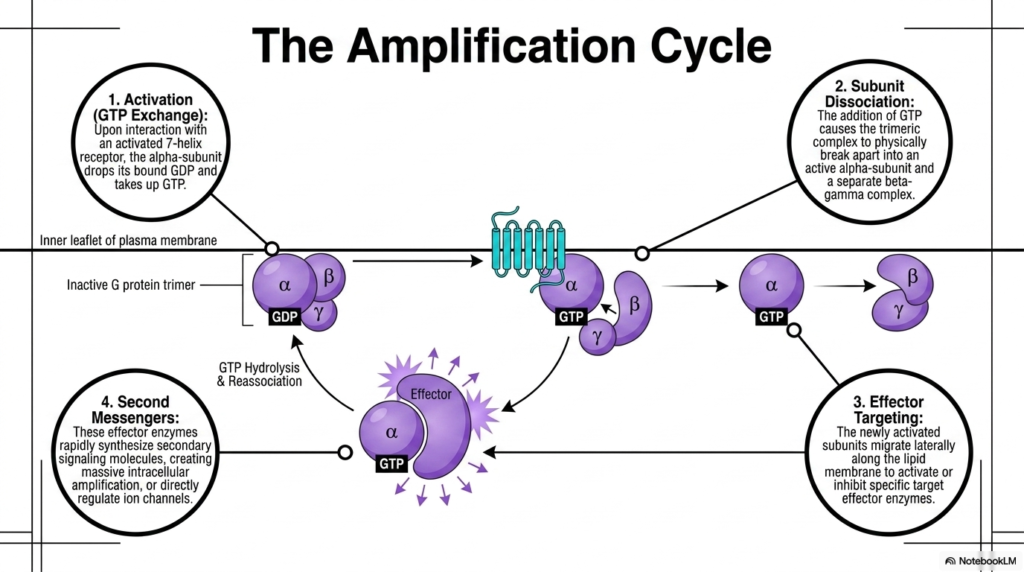
The ninth slide meticulously breaks down the dynamic Amplification Cycle, a hallmark feature of GPCR signaling pathways. The cycle initiates with Activation, specifically characterized by GTP Exchange. Upon direct physical interaction with newly activated 7-helix Membrane Receptors, the inactive G protein trimer undergoes a structural shift. This shift forces the alpha-subunit to forcefully drop its bound, low-energy GDP molecule and rapidly take up a high-energy GTP molecule from the surrounding cytoplasm. This simple nucleotide exchange is the definitive biochemical trigger that switches the G protein complex from a dormant state into a highly active state.
Following activation, the complex immediately undergoes Subunit Dissociation. The addition of the bulky, highly charged GTP molecule causes the once-stable trimeric complex to physically break apart. The structure splits into an independent, active alpha-subunit and a separate, combined beta-gamma complex. Both of these separated units remain anchored to the membrane, but they are now free to move independently. This dissociation step is crucial for Membrane Receptors, as it effectively multiplies the initial signal, creating two separate active entities that can independently hunt down and interact with different downstream cellular targets.
Step three involves Effector Targeting. The newly activated and separated subunits migrate rapidly laterally along the inner surface of the lipid membrane. Their goal is to locate and dock with specific target effector enzymes, either activating or inhibiting their function depending on the specific cellular context. Finally, step four results in the creation of Second Messengers. The targeted effector enzymes, once activated, quickly synthesize massive quantities of secondary signaling molecules. These Membrane Receptors thus orchestrate massive intracellular amplification, converting a single external activation event into thousands of internal second messengers that flood the cell.
To ensure the signal does not continue indefinitely, the alpha subunit has a built-in timer. It eventually hydrolyzes the attached GTP back into GDP, causing the entire G protein complex to reassociate and deactivate. This intrinsic reset mechanism is fundamental to maintaining proper physiological balance and preventing cellular overstimulation.
Slide 10: Immune Synapse Architectures and Specialized Membrane Receptors
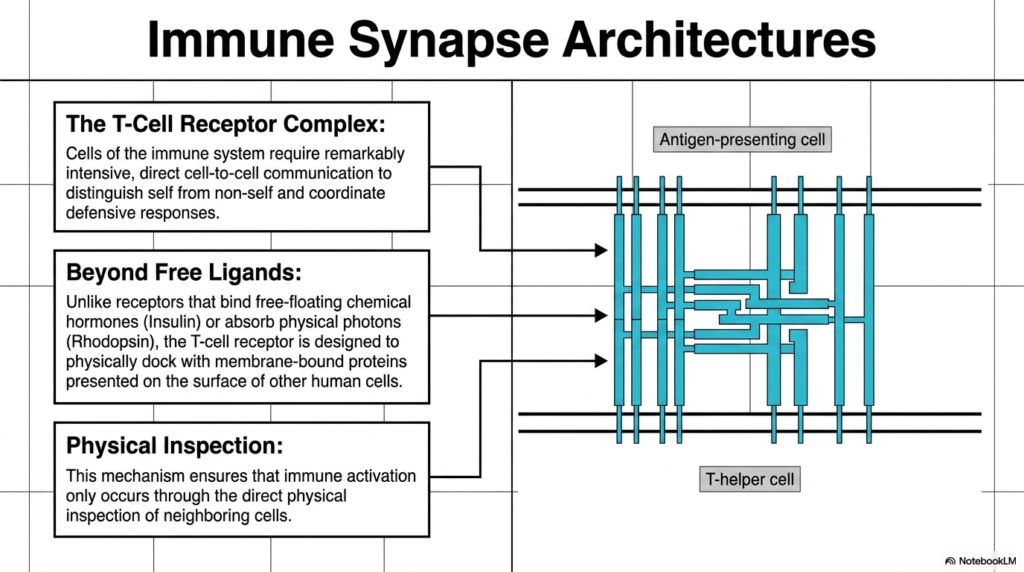
Moving into the realm of cellular defense, the tenth slide introduces the highly complex Immune Synapse Architectures, prominently featuring the T-Cell Receptor Complex. Cells of the immune system cannot afford errors; they require remarkably intensive, direct cell-to-cell communication to effectively distinguish friendly self-cells from dangerous non-self entities. This crucial defensive coordination is heavily reliant on specialized, multimeric Membrane Receptors. These complex protein structures allow immune cells to directly interrogate the surfaces of neighboring cells, acting as molecular security checkpoints that govern the incredibly destructive power of the human immune response.
The operational logic of the T-Cell Receptor goes far beyond interacting with simple free-floating ligands. Unlike the previously discussed Membrane Receptors that bind diffusing chemical hormones like insulin or absorb ephemeral physical photons like Rhodopsin, the T-cell receptor operates differently. It is explicitly designed and structurally engineered to physically dock with other rigid, membrane-bound proteins presented on the surfaces of other human cells. This requires the two cells to come into extremely close physical proximity, forming a tight junction that biologists refer to as an immune synapse.
This requirement for direct docking facilitates rigorous physical inspection. The T-cell uses these specialized Membrane Receptors to manually scan the molecular surface of the neighboring cell. This brilliant mechanism ensures that devastating immune activation occurs only through direct, confirmed physical inspection of neighboring tissues. By restricting activation to physical contact rather than distant chemical signals, the immune system prevents accidental, widespread inflammatory damage to healthy tissues, ensuring that the lethal immune response remains highly localized and entirely justified.
Understanding this synaptic architecture is a foundational principle for immunology students. The sheer complexity of multi-protein immune docks represents the pinnacle of cellular communication, highlighting how cells have evolved to construct elaborate physical bridges simply to safely exchange critical biological information regarding infection and cellular health.
Slide 11: Structural Recognition of Infection by Membrane Receptors
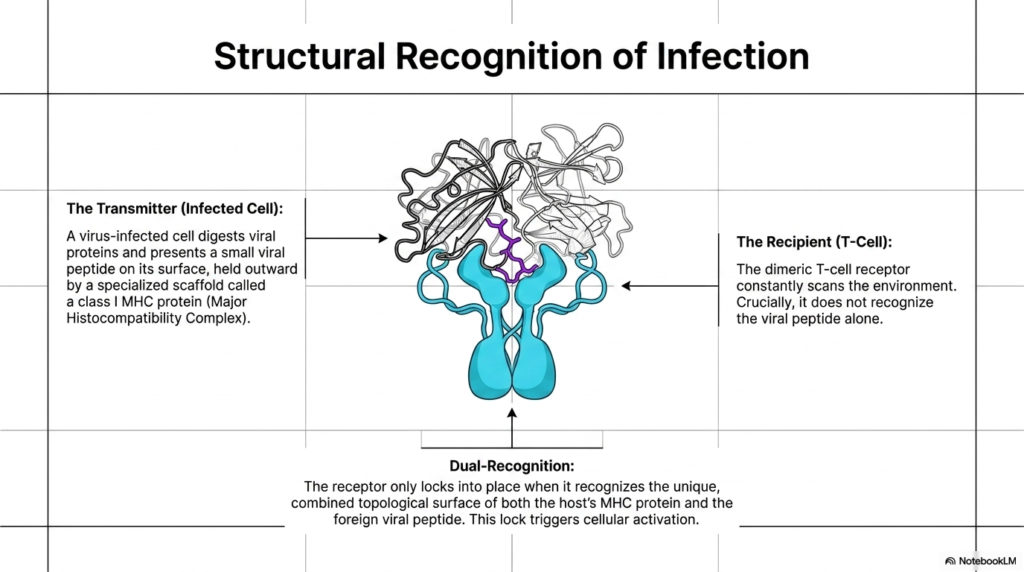
The eleventh slide dives into the precise molecular mechanism of this immune inspection, detailing the structural recognition of infection. The interaction occurs between a transmitter and a recipient. The transmitter is typically a virus-infected cell. When a cell becomes infected, it digests internal viral proteins and presents a tiny fragment, a small viral peptide, on its external surface. This peptide is held outward by a highly specialized cellular scaffold called a Class I MHC protein (Major Histocompatibility Complex). The infected cell uses this complex to wave a molecular red flag, signaling for help.
The recipient in this biological exchange is the patrolling T-Cell. As it navigates through tissues, it uses its dimeric T-cell receptor to constantly scan the surrounding environment. However, a crucial aspect of these specific Membrane Receptors is that they are structurally incapable of recognizing the isolated viral peptide floating alone. The receptor requires context. It must engage with the entire presentation scaffold to function correctly, preventing the immune system from blindly attacking harmless biological debris that may be drifting harmlessly through the extracellular fluid.
The activation of the T-cell relies on a stringent requirement known as Dual-Recognition. The dimeric T-cell receptor must simultaneously recognize both the host’s MHC protein structure and the foreign viral peptide nestled within it. Only when it recognizes this unique, combined topological surface will the receptor lock firmly into place. These specialized Membrane Receptors act like a highly complex keyhole that requires a two-part molecular key. This precise lock triggers a massive internal cellular activation cascade, commanding the T-cell to rapidly destroy the infected host cell to halt viral replication.
This dual-recognition paradigm is an incredible evolutionary safeguard. By demanding that Membrane Receptors verify both the identity of the host cell via the MHC and the presence of danger via the peptide, the body meticulously controls its most dangerous cellular weapons, a process that medical students must comprehend thoroughly.
Slide 12: A Structural Taxonomy of Membrane Receptors
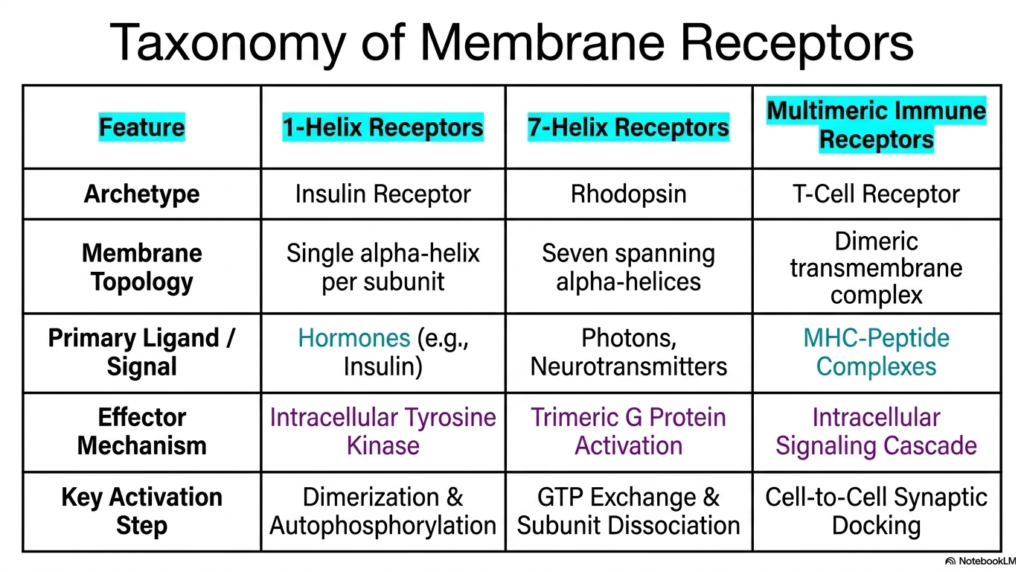
Synthesizing the diverse architectures explored thus far, the twelfth slide presents a comprehensive taxonomy of Membrane Receptors. This comparative table serves as a vital study tool, organizing these complex protein machines based on their distinguishing functional and structural features. The chart categorizes the cellular sensors into three primary classifications: 1-Helix Receptors, 7-Helix Receptors, and Multimeric Immune Receptors. By placing their distinct archetypes—the Insulin Receptor, Rhodopsin, and the T-Cell Receptor—side by side, students can easily visualize the evolutionary progression and the vast structural diversity that dictate modern cellular communication.
The table clearly delineates the membrane topology and the preferred primary signals for each category. For instance, the 1-helix models rely on a single alpha-helix per subunit and predominantly bind large, circulating chemical hormones like insulin. In stark contrast, the 7-helix models weave seven alpha-helical spans through the lipid bilayer and demonstrate incredible versatility, capturing everything from chemical neurotransmitters to physical photons. Meanwhile, the multimeric immune complexes utilize massive dimeric transmembrane structures specifically to engage with complex, membrane-bound MHC-peptide signals. These distinct topological features fundamentally dictate what kinds of messages these Membrane Receptors can interpret.
Furthermore, the taxonomy highlights the drastically different effector mechanisms and key activation steps utilized by each class. 1-helix systems utilize built-in intracellular tyrosine kinases driven by dimerization and autophosphorylation. Conversely, 7-helix systems rely entirely on external trimeric G protein activation facilitated by complex GTP exchange and subunit dissociation. Finally, immune structures rely on direct cell-to-cell synaptic docking to trigger specialized intracellular signaling cascades. Understanding this taxonomy of Membrane Receptors provides a foundational framework, allowing scholars to predict exactly how a newly discovered biological signal will likely alter cellular behavior based simply on receptor architecture.
By categorizing these profound molecular mechanisms side by side, this table distills immense biochemical complexity into an accessible format. Mastering this comparative chart ensures that aspiring medical professionals can confidently navigate the complex web of pharmacology, cellular biology, and modern immunology.
Slide 13: The Universal Grammar of Signal Transduction via Membrane Receptors
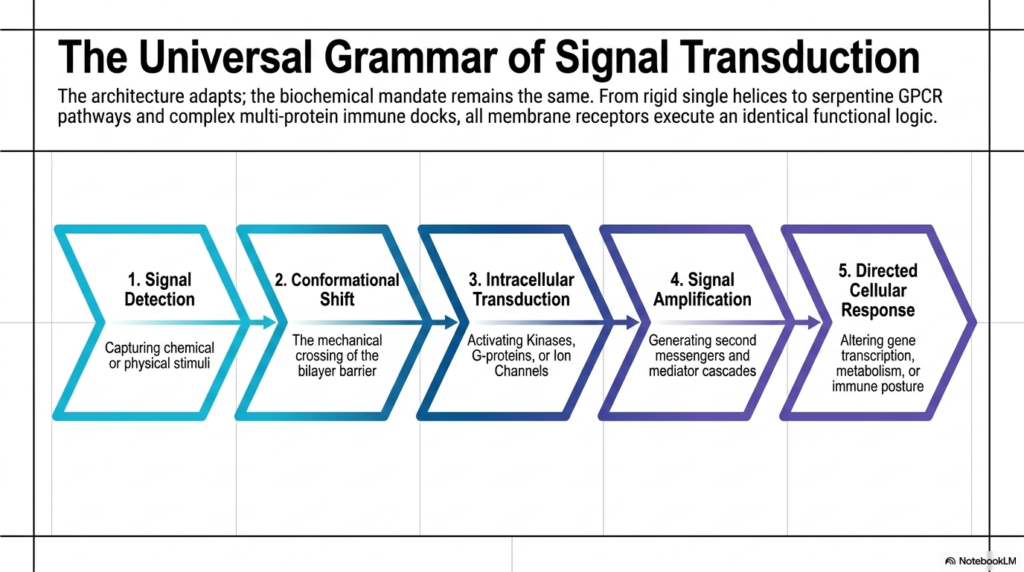
The final slide brilliantly summarizes the entire presentation by establishing a universal grammar of signal transduction. While the physical architectures adapt incredibly across different tissues and organisms, the underlying biochemical mandate remains remarkably consistent. Whether a cell is using rigid single helices, complex serpentine GPCR pathways, or massive multi-protein immune docks, all Membrane Receptors ultimately follow identical, step-by-step functional logic. This underlying logical sequence unites the vast, seemingly chaotic diversity of cellular biology into a highly predictable and understandable five-step biological framework that governs all cellular life.
This universal grammatical sequence always begins with Signal Detection, in which the receptor captures specific chemical or physical stimuli from the extracellular space. This is immediately followed by a Conformational Shift, representing the mechanical crossing of the lipid bilayer barrier. The third crucial phase is Intracellular Transduction. Here, the internal domains of the Membrane Receptors spring into action, activating specific kinases, liberating G-proteins, or forcefully opening internal ion channels. This step marks the true transfer of information from outside the cell into the protected internal cytoplasm, converting physical movement into biological chemistry.
The sequence continues rapidly into phase four: Signal Amplification. Through the robust generation of abundant second messengers and cascading mediator pathways, a tiny, singular external event is exponentially magnified inside the cell. Finally, this massive amplification culminates in the fifth phase: a Directed Cellular Response. The original message, received and translated by the Membrane Receptors, ultimately succeeds in altering core gene transcription, fundamentally shifting cellular metabolism, or adopting a highly aggressive immune posture. This predictable five-step logic underscores the beautiful simplicity hidden within the immense complexity of cellular physiology.
By recognizing this universal functional sequence, students of biochemistry are no longer forced to memorize isolated, disconnected facts. Instead, they can confidently apply this logical framework to any newly discovered pathway, truly mastering the elegant language of cellular communication and its impact on human health.
Please read our Content Disclaimer Statement.
Check out our social media channels: