77. Protein Sorting and Exocytosis: Cellular Mechanisms
Imagine a bustling city without addresses or delivery trucks; chaos would ensue. Cells face a similar logistical challenge. They produce millions of proteins that must reach precise locations to function. This slide deck explores the molecular routing systems that solve this problem. Understanding these mechanisms reveals how life maintains order at a microscopic level, ensuring survival and health.
Slide 1: Welcome to Protein Sorting and Exocytosis
The journey of a protein from its creation to its final destination is a marvel of biological engineering. At the heart of this complex logistical network lies the dual phenomenon of Protein Sorting and Exocytosis. This initial slide introduces the fundamental concept that cells possess highly regulated biochemical pathways and cellular routing mechanisms. These microscopic highways ensure that every synthesized molecule reaches its proper functional location. Without Protein Sorting and Exocytosis, a cell would quickly accumulate misplaced enzymes and structural components, leading to catastrophic metabolic failure and cell death.
Consider the sheer volume of material a single cell processes every minute. The first slide visually represents a secretory vesicle fusing with the plasma membrane, a hallmark event. This visual perfectly captures the essence of Protein Sorting and Exocytosis, showcasing how bulk materials are expelled from the cell into the extracellular environment. The downward-pointing arrow indicates the direction of flow for these critical biochemical products. It provides a simplified snapshot of a highly intricate, energy-dependent process that keeps complex tissues and entire organisms functioning smoothly.
Understanding these cellular pathways is vital for modern medical and biological sciences. The complex mechanisms governing Protein Sorting and Exocytosis are directly implicated in numerous vital physiological responses, ranging from insulin secretion in the pancreas to neurotransmission in the brain. When these pathways operate correctly, organisms maintain homeostasis and thrive in their environments. However, slight molecular errors in these delicate routing protocols can cause severe diseases.
Conditions such as cystic fibrosis, various neurodegenerative disorders, and diverse metabolic syndromes often originate from defects in these very mechanisms. Therefore, mastering these foundational biochemical concepts is absolutely essential. For college and medical students, recognizing how molecules navigate the intracellular space provides the groundwork for understanding both basic physiology and complex pharmacological interventions aimed at correcting cellular misrouting.
Slide 2: Origin Pathways in Protein Sorting and Exocytosis
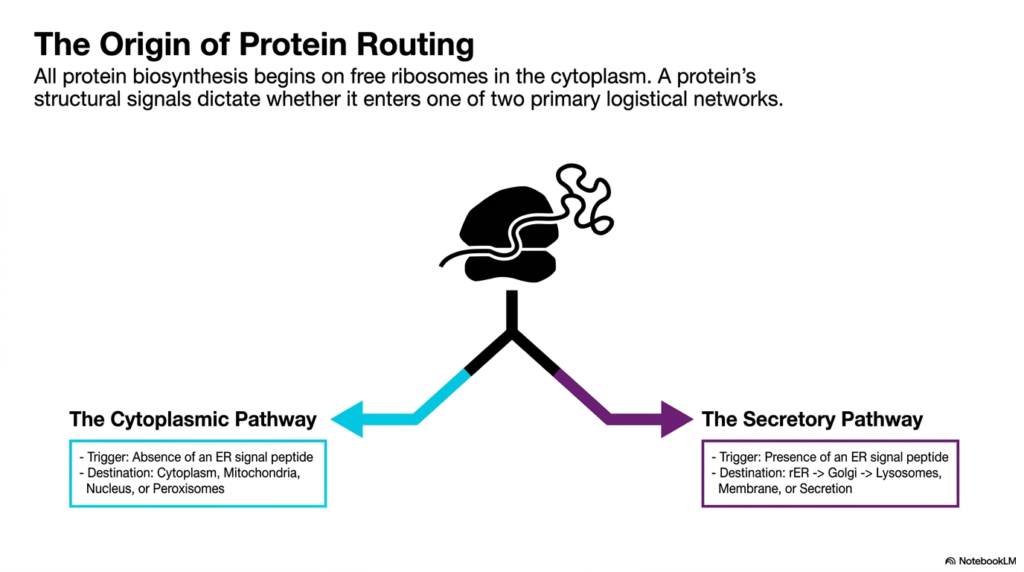
Every functional protein begins its existence in the cytoplasm, where it is translated by free ribosomes. The second slide illustrates the critical intersection where a nascent polypeptide’s fate is decided. This crucial branching point is foundational to understanding Protein Sorting and Exocytosis. The presence or absence of specific structural signals embedded within the protein determines whether it enters one of two primary logistical networks. This decision dictates the entire future trajectory of the molecule, separating those destined for internal housekeeping from those meant for the secretory apparatus.
The diagram clearly delineates the Cytoplasmic Pathway from the Secretory Pathway. If a newly synthesized protein lacks a specific Endoplasmic Reticulum signal peptide, it defaults to the Cytoplasmic Pathway. These proteins generally remain in the cytoplasm or travel to destinations such as mitochondria, the nucleus, or peroxisomes. Conversely, the presence of an Endoplasmic Reticulum signal peptide acts as a definitive trigger, shunting the protein into the Secretory Pathway. This sorting step is a master regulator of cellular organization.
Entering the Secretory Pathway is the first step toward the extracellular space. The graphic outlines the typical route: from the rough Endoplasmic Reticulum to the Golgi apparatus, and finally toward lysosomes, the membrane, or secretion. This is where Protein Sorting and Exocytosis truly intertwine with cellular communication. By strictly controlling which proteins enter this pathway, the cell ensures that only appropriate molecules, such as hormones or digestive enzymes, are prepared for eventual release.
The elegance of this system lies in its inherent simplicity at the decision point. A single molecular tag dictates a complex biochemical cascade. Understanding this initial divergence is paramount for medical students, as it highlights how genetic mutations altering a signal sequence can misdirect a protein entirely. Such misdirection disrupts Protein Sorting and Exocytosis, triggering a cascade of cellular dysfunction and underscoring the critical importance of primary amino acid sequences in overall biological processes.
Slide 3: Organelle Transit in Protein Sorting and Exocytosis
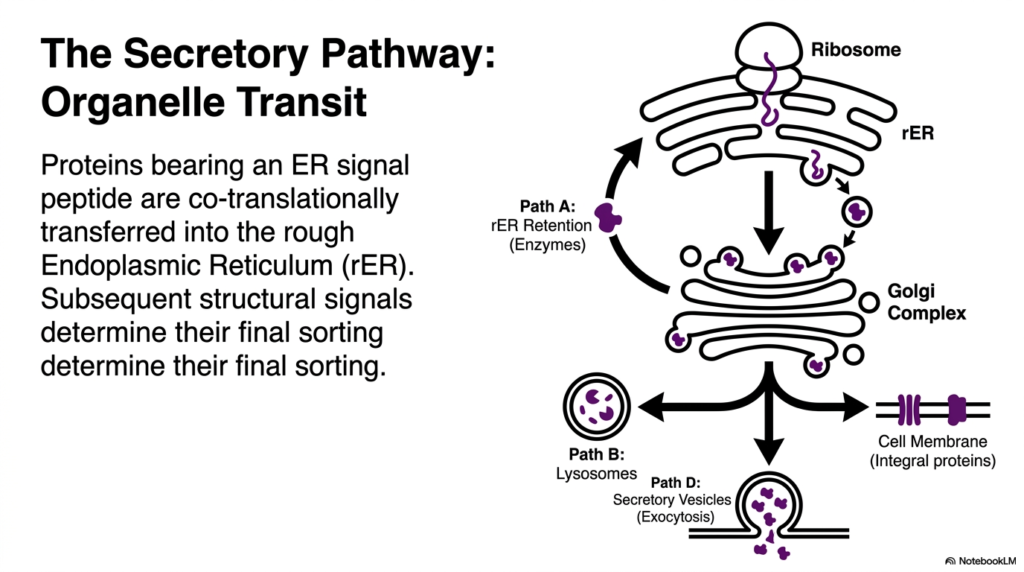
Once a protein is committed to the Secretory Pathway, it begins a complex journey through the cell’s internal organelles. This slide maps the transit routes that are central to Protein Sorting and Exocytosis. Proteins possessing an Endoplasmic Reticulum signal peptide are co-translationally transferred directly into the rough Endoplasmic Reticulum. Here, the protein begins folding into its three-dimensional conformation while undergoing crucial initial modifications. Subsequent structural signals within the polypeptide chain then determine its final sorting destination among several distinct pathways.
The visual brilliantly breaks down the four primary routes branching from the Golgi complex, a major hub for Protein Sorting and Exocytosis. Path A represents retention within the rough Endoplasmic Reticulum, typically reserved for essential resident enzymes needed for metabolic functions. Path B shows sorting to lysosomes, directing degradative enzymes to the cell’s waste disposal units. These internal sorting decisions highlight the intricate sorting capacity of the Golgi network, which acts much like a cellular post office analyzing chemical zip codes.
Path D is perhaps the most dynamic, representing the culmination of the secretory route. Here, proteins are packaged into secretory vesicles destined for the cell membrane. This specific route is the direct precursor to the final fusion event, embodying the core action of Protein Sorting and Exocytosis. The vesicles travel toward the periphery, carrying cargo like neurotransmitters or hormones. Once they reach the boundary, they release their contents outward, a vital process for intercellular signaling and systemic physiological responses.
Finally, the diagram illustrates the pathway for integral membrane proteins. Instead of being released, these proteins become permanently embedded in the cell membrane. Each of these paths requires precise molecular recognition and vesicular transport mechanisms. For students of biochemistry, dissecting these distinct transit routes reveals the sheer complexity required to maintain cellular equilibrium. It demonstrates that every protein’s journey is meticulously calculated and executed in the context of Protein Sorting and Exocytosis.
Slide 4: Direct Targeting in Protein Sorting and Exocytosis
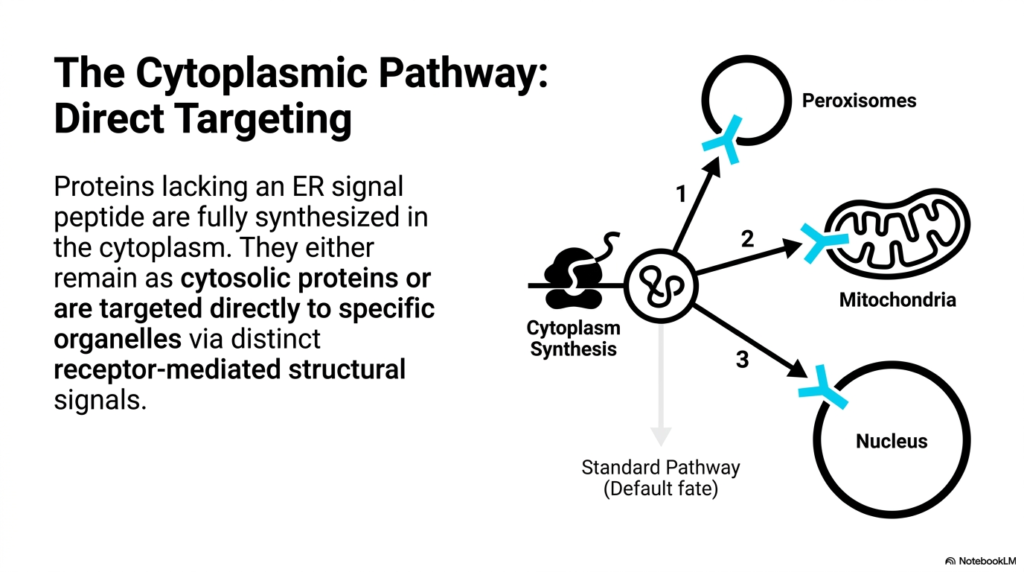
Not all proteins are destined for the complex vesicular networks of the secretory system. This slide focuses on the Cytoplasmic Pathway, which runs parallel to the mechanisms of Protein Sorting and Exocytosis. Proteins that lack an Endoplasmic Reticulum signal peptide are fully synthesized on free ribosomes directly within the cytoplasm. This pathway represents a separate, yet equally vital, logistical network dedicated to supplying the cell’s internal organelles with the specific biochemical machinery they require to function properly and maintain energy production.
Unlike the vesicular transport seen in the secretory route, the Cytoplasmic Pathway relies on direct targeting. Proteins synthesized here typically adopt a default fate of remaining as cytosolic proteins unless they possess distinct receptor-mediated structural signals. The slide illustrates the specific routes to three major organelles: the peroxisomes, mitochondria, and the nucleus. Each numbered pathway represents a highly selective import process, ensuring that the cytoplasm does not become depleted of its essential structural and functional components.
The contrast between these two major cellular routes is a fascinating aspect of biology. While Protein Sorting and Exocytosis focuses largely on packaging and export or membrane integration, the Cytoplasmic Pathway is about direct, targeted delivery across organelle membranes. For instance, mitochondrial proteins must unfold to thread through transport complexes in the double membrane. Understanding these distinct import mechanisms is crucial for comprehending how cells sustain their internal metabolism independently of their external secretory obligations.
Recognizing the division of labor between these pathways provides a holistic view of cell biology. While researchers often highlight Protein Sorting and Exocytosis for their role in systemic communication, the Cytoplasmic Pathway is the bedrock of cellular survival. Medical students must appreciate that defects in these direct targeting mechanisms can lead to severe mitochondrial or peroxisomal disorders. Such pathologies highlight the absolute necessity of precise intracellular protein routing to maintain cellular health and prevent metabolic collapse.
Slide 5: Translocation Signals in Protein Sorting and Exocytosis
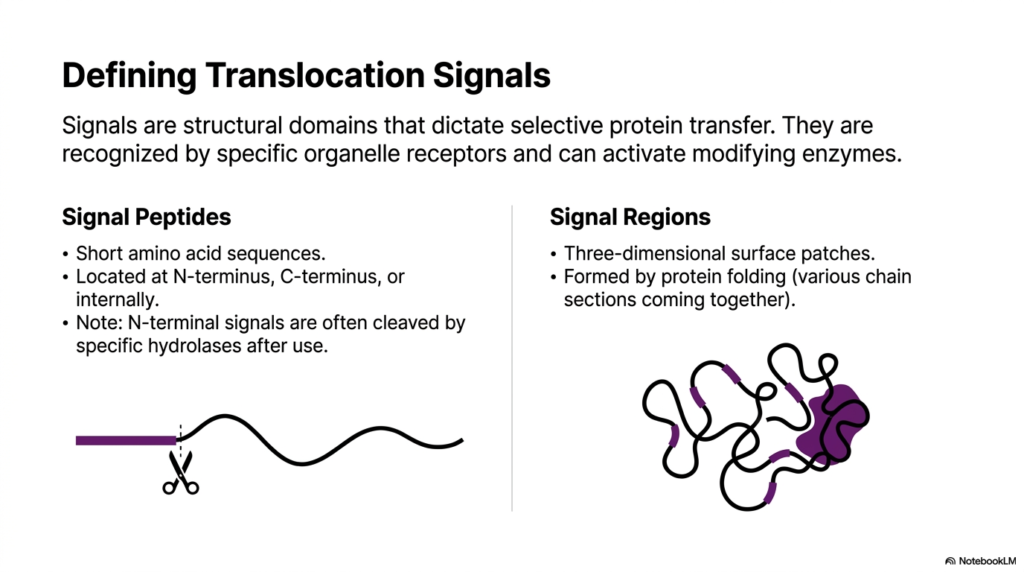
The precise navigation of a protein depends entirely on its built-in molecular addresses. This slide defines these critical translocation signals, which are the fundamental vocabulary of Protein Sorting and Exocytosis. Signals are essentially structural domains that dictate selective protein transfer across cellular compartments. They are recognized by specific organelle receptors, triggering transport mechanisms or activating modifying enzymes. Without these precise chemical tags, proteins would drift aimlessly, completely breaking down the necessary order of the cellular environment.
The slide differentiates between two primary types of addressing tags: Signal Peptides and Signal Regions. Signal Peptides are short, linear sequences of amino acids typically located at the N-terminus, C-terminus, or occasionally internally. Notably, N-terminal signals are often cleaved by specific hydrolases immediately after they fulfill their routing function. This cleavage is a defining characteristic of early steps in Protein Sorting and Exocytosis, ensuring that the transport tag does not interfere with the mature protein’s final biochemical structure or its enzymatic activity.
In contrast, Signal Regions rely on three-dimensional protein folding rather than a simple linear sequence. As the linear amino acid chain folds, various distant sections of the chain are brought together in three-dimensional space to form a unique surface patch. This structural patch acts as the signal. Understanding this distinction is vital for biochemists, as it illustrates that cellular receptors can read both simple linear codes and complex topographical features when determining a protein’s proper destination.
Mastering the concept of these signals is central to understanding Protein Sorting and Exocytosis. These molecular addresses are the keys that unlock specific cellular compartments. For medical students, recognizing how these signals function opens the door to understanding molecular medicine. Many advanced therapeutics and viral hijacking mechanisms exploit these exact translocation signals. By mimicking or altering these patches and peptides, one can theoretically redirect cellular traffic to treat various metabolic and genetic disorders directly affecting Protein Sorting and Exocytosis.
Slide 6: ER Entry Mechanisms in Protein Sorting and Exocytosis
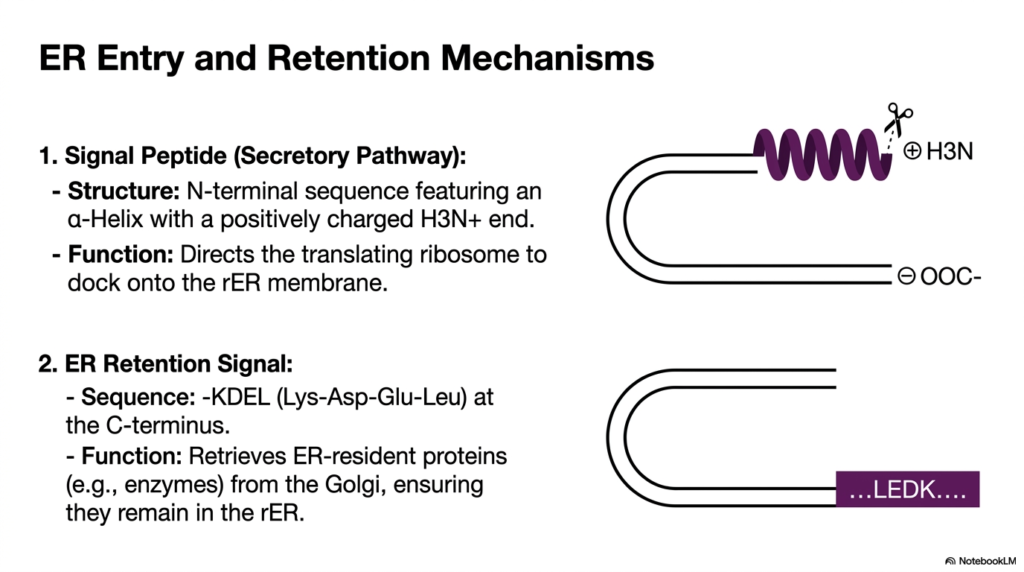
The rough Endoplasmic Reticulum serves as the primary gateway for the secretory route. This slide details the specific molecular interactions required for ER entry and retention, foundational events in Protein Sorting and Exocytosis. The first mechanism shown is the Secretory Pathway Signal Peptide. This structure is defined by an N-terminal sequence featuring a distinct alpha-helix capped with a positively charged end. Its primary function is to direct the actively translating ribosome to physically dock onto the membrane of the rough ER.
This docking event represents a brilliant biochemical adaptation. By tethering the ribosome to the membrane, the cell ensures that the newly forming polypeptide is threaded directly into the protected ER lumen as it is synthesized. This co-translational translocation prevents hydrophobic regions of the new protein from clumping improperly in the aqueous cytoplasm. It is a critical quality-control checkpoint in Protein Sorting and Exocytosis, allowing for immediate modification and proper folding by resident ER chaperones.
The slide also highlights a second crucial mechanism: the ER Retention Signal. Certain proteins, such as folding enzymes, must remain in the ER to function. They are marked by a specific KDEL sequence at their C-terminus. Interestingly, these proteins frequently escape to the Golgi apparatus during bulk transport. The KDEL signal functions as a retrieval mechanism. Receptors in the Golgi recognize this sequence and actively shuttle the escaped resident proteins back to the rough ER, maintaining organelle identity.
This continuous retrieval loop perfectly illustrates the dynamic nature of Protein Sorting and Exocytosis. It is not just about moving things forward; it involves meticulous recycling and error correction. Medical professionals must understand these retention mechanisms because disruptions can lead to a depletion of essential ER enzymes. Such depletion leads to the accumulation of misfolded proteins, triggering cellular stress responses that are deeply implicated in numerous inflammatory and neurodegenerative disease states linked to errors in Protein Sorting and Exocytosis.
Slide 7: Lysosomal Addressing in Protein Sorting and Exocytosis
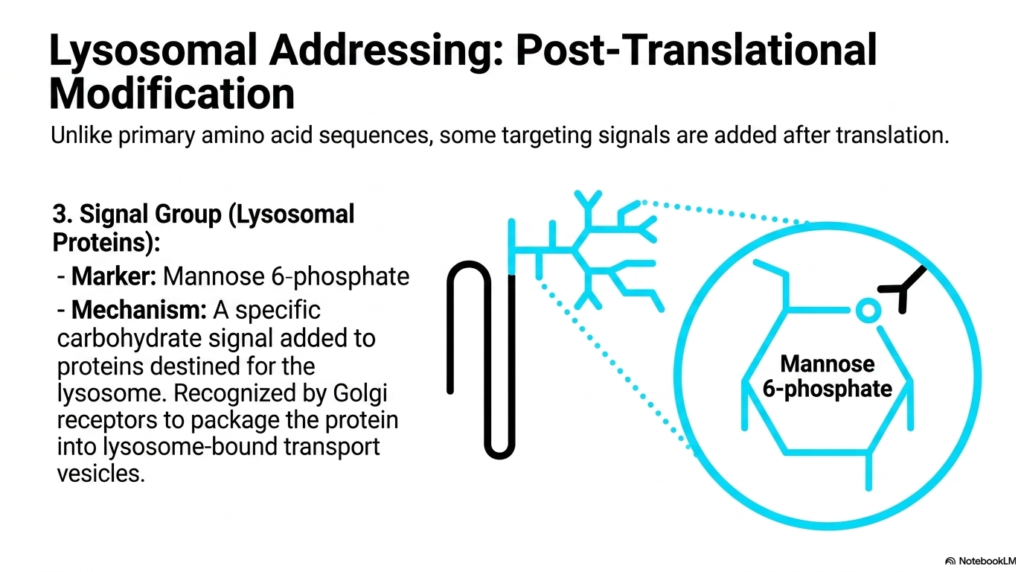
Not all molecular addresses are written directly into a protein’s primary DNA sequence. This slide introduces the concept of post-translational modification in routing, a highly specialized branch of Protein Sorting and Exocytosis. The focus here is on Lysosomal Addressing, which relies on a uniquely added signal group rather than a standard amino acid peptide. The specific marker is Mannose 6-phosphate, a carbohydrate molecular tag attached to proteins specifically destined for the cell’s lysosome, the primary degradative organelle.
Unlike primary amino acid sequences translated directly from mRNA, this carbohydrate signal is enzymatically added after the protein has already entered the secretory pathway. The detailed chemical structure of Mannose 6-phosphate is shown, highlighting its specific conformation. This mechanism represents an advanced layer of complexity in Protein Sorting and Exocytosis. It demonstrates how cells utilize complex sugar modifications, known as glycosylation, to encode routing information, expanding the cell’s logistical capabilities beyond simple peptide-based recognition systems.
Once the Mannose 6-phosphate tag is securely attached in the Golgi apparatus, it acts as a precise chemical beacon. Specific Golgi receptors bind tightly to this carbohydrate marker, ensuring that these dangerous degradative enzymes are safely segregated. They are then packaged into specialized transport vesicles bound strictly for the lysosome. This segregation is paramount; if these enzymes were mistakenly routed to the cell membrane or cytoplasm, they would begin digesting the cell’s own structural components, leading to rapid self-destruction.
Understanding this post-translational modification is a major milestone for students studying Protein Sorting and Exocytosis. It provides a clear mechanical basis for devastating conditions known as lysosomal storage disorders. In diseases like I-cell disease, the enzyme responsible for adding the Mannose 6-phosphate tag is defective. Consequently, crucial lysosomal proteins are secreted outside the cell rather than reaching the lysosome, thereby demonstrating what happens when this intricate molecular addressing system in Protein Sorting and Exocytosis fails.
Slide 8: Membrane Integration in Protein Sorting and Exocytosis
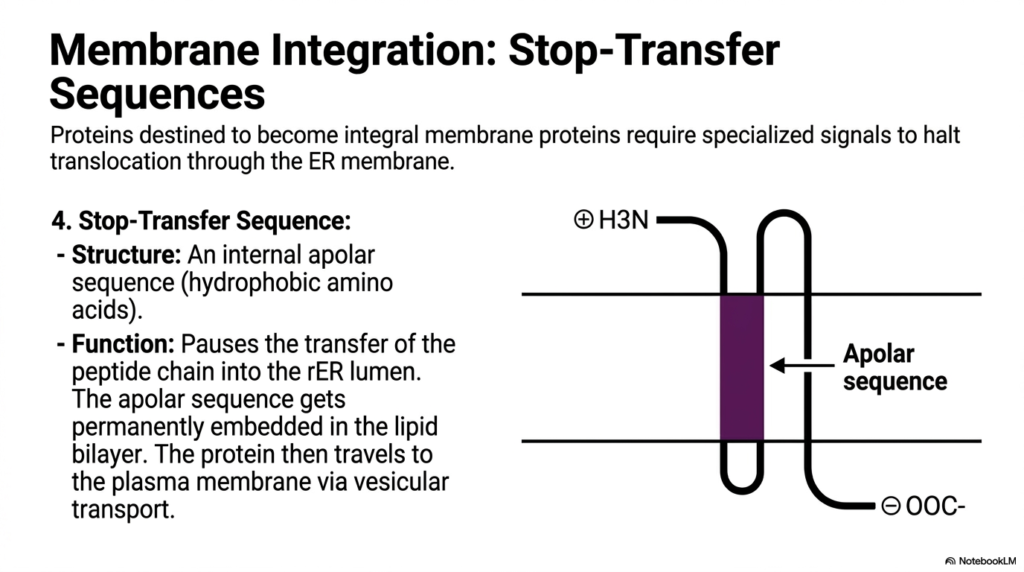
A crucial function of cellular logistics is the construction of the cell’s outer boundary. This slide explains how proteins destined to become integral membrane proteins are embedded, a process closely linked to Protein Sorting and Exocytosis. These specific proteins require specialized signals to halt their forward translocation through the Endoplasmic Reticulum membrane. Without these specific internal signals, the protein would be pushed entirely into the ER lumen, ultimately becoming a free-floating soluble protein designated for secretion rather than membrane integration.
The key to this process is the Stop-Transfer Sequence. Structurally, this is an internal apolar sequence composed heavily of hydrophobic amino acids. As the nascent polypeptide threads through the ER membrane channel, this hydrophobic stretch acts as a physical brake. It pauses the transfer of the peptide chain, causing the transport machinery to release the protein laterally. Consequently, the apolar sequence becomes permanently embedded directly within the hydrophobic core of the lipid bilayer, anchoring the protein securely in place.
Once properly anchored in the ER membrane, these integral proteins are not static. The surrounding membrane buds off to form transport vesicles. This vesicular transport is a core mechanical feature of Protein Sorting and Exocytosis. The vesicles, carrying the newly integrated proteins, travel to the Golgi and eventually to the plasma membrane. When the vesicle fuses with the outer cell boundary, the embedded proteins become permanent fixtures of the cell’s exterior surface, functioning as crucial receptors or ion channels.
For medical students, visualizing this integration process is essential for understanding cellular physiology. Almost all pharmacological drugs target integral membrane proteins, such as G protein-coupled receptors. Understanding how the Stop-Transfer Sequence functions during Protein Sorting and Exocytosis explains how these massive receptor complexes are initially built and deployed. Anomalies in this embedding process can lead to the omission of cellular receptors, resulting in profound communication failures between tissues and halting Protein Sorting and Exocytosis.
Slide 9: Cytoplasmic Addressing in Protein Sorting and Exocytosis
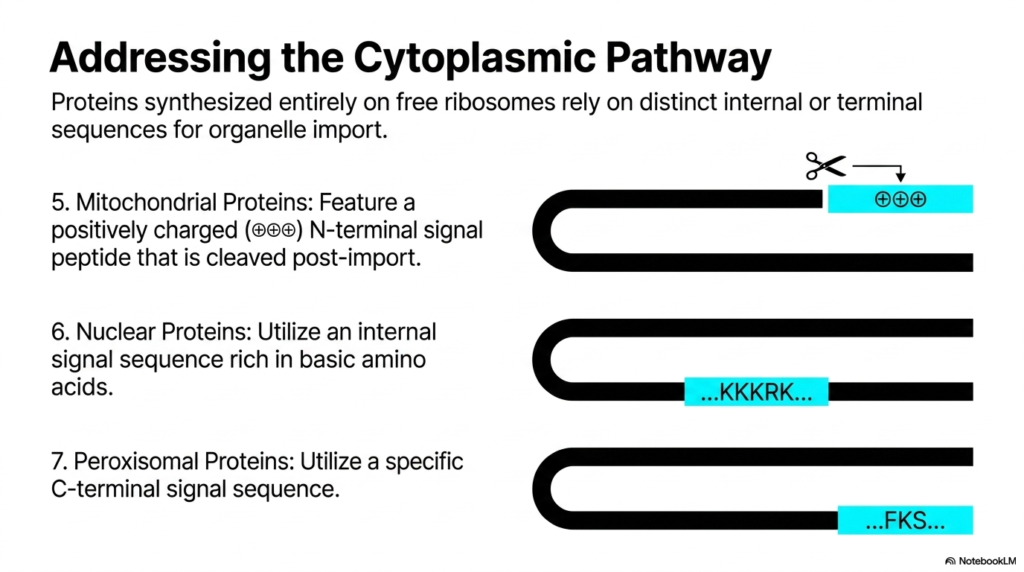
Returning to the alternative route, this slide provides the specific molecular codes used for direct organelle import. While distinct from the vesicular transport of Protein Sorting and Exocytosis, understanding these signals completes the cellular logistics picture. Proteins synthesized entirely on free ribosomes rely on distinct internal or terminal sequences for their import into organelles. This mechanism bypasses the Endoplasmic Reticulum entirely, using specialized translocator channels on the target organelle’s outer membrane to achieve precise intracellular delivery.
The slide outlines three distinct addressing codes. Mitochondrial proteins feature a positively charged N-terminal signal peptide. Interestingly, similar to some secretory signals, this peptide is cleaved post-import by resident proteases. Nuclear proteins use a completely different strategy: they rely on an internal signal sequence rich in basic amino acids, commonly denoted KKKRK. This allows fully folded proteins to pass through the massive nuclear pore complexes without structural unfolding.
Finally, the diagram highlights Peroxisomal proteins, which utilize a specific C-terminal signal sequence for their targeting. The diversity of these codes showcases the evolutionary complexity of cellular logistics. While Protein Sorting and Exocytosis handle the bulk export and membrane construction, these distinct cytoplasmic signals maintain the internal power plants and genetic vaults. Each organelle requires a unique chemical lock-and-key mechanism to prevent inappropriate proteins from entering and disrupting highly specialized, isolated biochemical microenvironments.
Understanding these alternative routing mechanisms is vital for a comprehensive biochemical education. Although not strictly part of Protein Sorting and Exocytosis, these pathways share the fundamental principle of signal-based targeting. Medical students must recognize these distinct sequences because mutations at these sites lead to specific, often fatal, organelle-centric diseases. For instance, defective peroxisomal targeting sequences result in devastating neurological conditions, illustrating that proper internal routing is just as critical as proper execution of Protein Sorting and Exocytosis.
Slide 10: The Machinery of Protein Sorting and Exocytosis
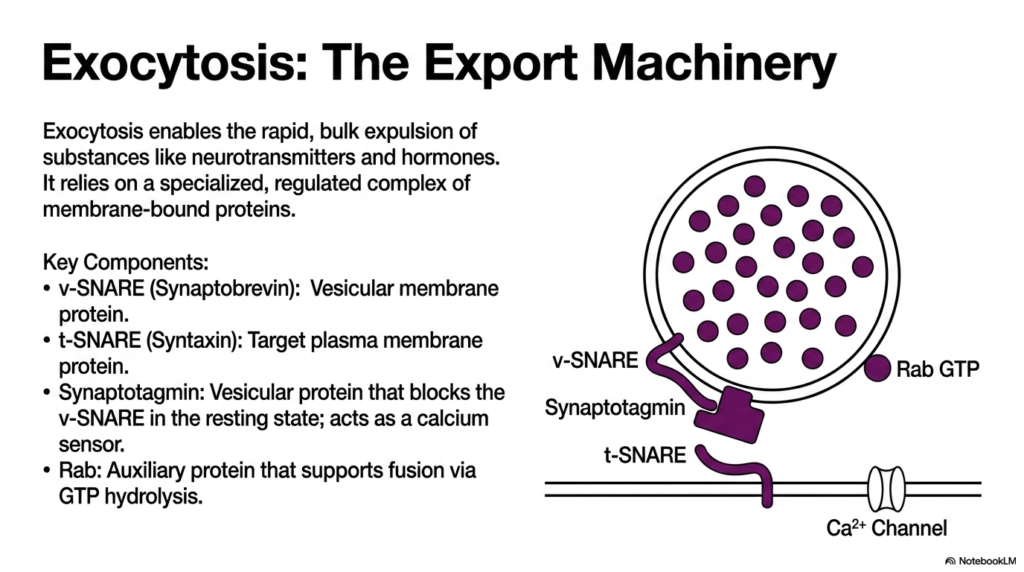
We now reach the terminal event of the secretory pathway. This slide details the sophisticated export machinery that drives the final stages of Protein Sorting and Exocytosis. Exocytosis enables the rapid, bulk expulsion of vital substances like neurotransmitters and metabolic hormones into the extracellular space. This is not a passive leaking process; it relies on a highly specialized, tightly regulated complex of membrane-bound proteins that work in concert to physically force two repelling lipid bilayers to merge.
The graphic introduces the critical components of the SNARE complex. The v-SNARE, or Synaptobrevin, is a vital vesicular membrane protein. Its counterpart is the t-SNARE, or Syntaxin, which is anchored securely to the target plasma membrane. In a resting state, this system is primed but halted. A key vesicular protein called Synaptotagmin acts as a calcium sensor. It actively blocks the v-SNARE, preventing premature fusion and ensuring that the final step of Protein Sorting and Exocytosis only occurs when a specific physiological command is received.
Additionally, the process uses an auxiliary protein called Rab. Rab is a GTPase that supports fusion by hydrolyzing GTP, providing the necessary regulation and energy. The interplay between these specific proteins constitutes the core molecular engine of Protein Sorting and Exocytosis. The SNARE proteins act like a biochemical zipper, physically pulling the vesicle and cell membrane close enough to overcome electrostatic repulsion, a magnificent feat of microscopic mechanical engineering.
For medical students, the SNARE complex is a high-yield topic. This exact molecular machinery is what allows a neuron to fire and a muscle to contract. The precision of Protein Sorting and Exocytosis at this stage is absolute; a delay of even a millisecond in this fusion event would severely disrupt neural communication. Understanding the identities and resting states of v-SNAREs and t-SNAREs is foundational for grasping both normal neurophysiology and the mechanisms of various potent neurotoxins that target Protein Sorting and Exocytosis.
Slide 11: The Step-by-Step Process of Protein Sorting and Exocytosis
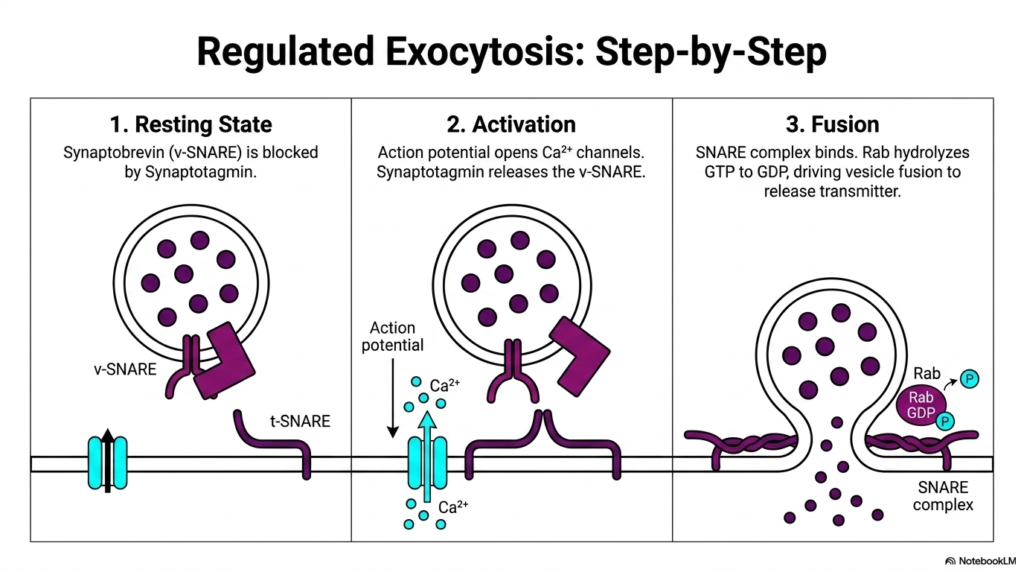
Building upon the previous components, this slide animates the dynamic sequence of regulated secretion. It breaks down the ultimate event in Protein Sorting and Exocytosis into three distinct, highly choreographed steps. In Step 1, the Resting State, the vesicle is fully loaded and positioned near the membrane. The v-SNARE is physically blocked by Synaptotagmin, maintaining a critical holding pattern. The cell is primed, holding its cargo until the precise moment a physiological signal dictates release.
Step 2 illustrates the Activation phase, the turning point of Protein Sorting and Exocytosis. An action potential reaches the membrane, causing voltage-gated calcium channels to snap open. The influx of calcium ions represents the biological trigger. Calcium binds rapidly to Synaptotagmin. This binding alters Synaptotagmin’s conformation, causing it to release its inhibitory grip on the v-SNARE. This calcium-dependent activation beautifully demonstrates how electrical signals in the nervous system are directly translated into mechanical and biochemical actions at the cellular level.
The final sequence, Step 3, is Fusion. With the block removed, the v-SNARE and t-SNARE complex rapidly bind together, twisting into a tight bundle that violently pulls the vesicle into the plasma membrane. Concurrently, the Rab protein hydrolyzes its bound GTP to GDP, thereby driving vesicle fusion. This mechanical zipping forces the lipid bilayers to mix, completely opening the vesicle pore and rapidly expelling the neurotransmitters into the synaptic cleft, completing the grand process of Protein Sorting and Exocytosis.
This step-by-step breakdown is crucial for students analyzing synaptic transmission. The mechanisms of Protein Sorting and Exocytosis detailed here are the very foundation of human movement, thought, and glandular secretion. Medical pharmacology heavily relies on understanding these three steps. By understanding exactly how calcium influx triggers the SNARE complex, researchers can develop sophisticated drugs that either enhance or inhibit Protein Sorting and Exocytosis, offering treatments for conditions ranging from severe muscle spasms to complex psychiatric disorders.
Slide 12: Pathological Disruption in Protein Sorting and Exocytosis
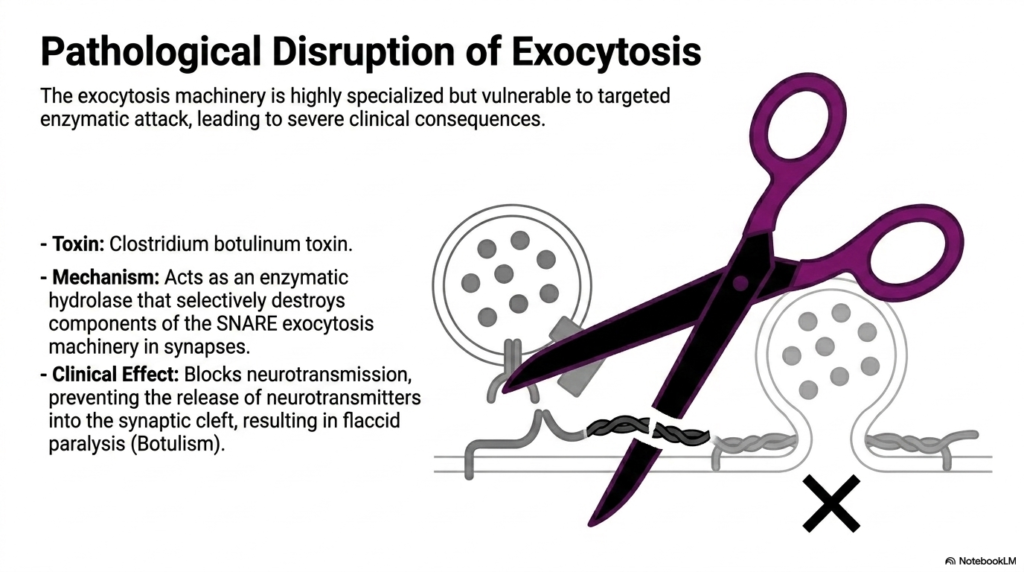
Even the most sophisticated biological machinery has vulnerabilities. This slide examines the severe clinical consequences that occur when the mechanisms of Protein Sorting and Exocytosis are subjected to pathological disruption. While the exocytosis machinery is highly specialized and generally robust, it is incredibly vulnerable to targeted enzymatic attacks by specific environmental toxins. The visual metaphor of scissors cutting the SNARE complex perfectly encapsulates the devastating, precise nature of these biological weapons on cellular infrastructure.
The specific pathology highlighted here involves the infamous Clostridium botulinum toxin. The mechanism of action is brilliantly sinister; the toxin acts as a highly specific enzymatic hydrolase. Once inside the cell, it selectively targets and destroys the protein components of the SNARE exocytic machinery at neural synapses. By physically cleaving the v-SNAREs or t-SNAREs, the toxin irreversibly breaks the essential zipper mechanism. This targeted destruction represents a complete, catastrophic failure of Protein Sorting and Exocytosis at the critical terminal stage.
The clinical effect of this molecular disruption is profound. Because the SNARE complex is disrupted, vesicle fusion is impossible. This completely blocks neurotransmission, preventing the release of vital neurotransmitters like acetylcholine into the synaptic cleft. The systemic result of this microscopic failure in Protein Sorting and Exocytosis is flaccid paralysis, the hallmark symptom of Botulism. Without the ability to release neurotransmitters, muscles cannot contract, ultimately leading to fatal respiratory failure if left untreated.
Studying this toxin offers medical students a profound lesson in applied biochemistry. It demonstrates that Protein Sorting and exocytosis are not just abstract cellular biology; they are the literal mechanism of survival. Understanding how botulinum toxin dismantles the export machinery provides a perfect, real-world application of the molecular components learned in earlier slides. Furthermore, it explains the biochemical rationale behind using controlled, minute doses of this same toxin clinically, such as Botox, to selectively manage severe muscle spasticity by regulating Protein Sorting and Exocytosis.
Slide 13: The Master Matrix of Protein Sorting and Exocytosis
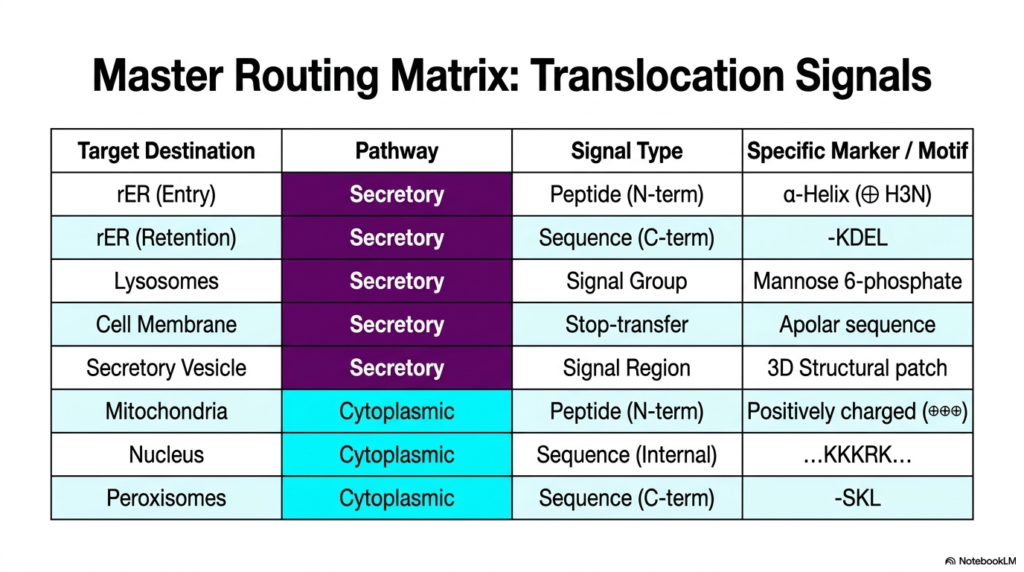
This final slide serves as a comprehensive synthesis of the entire lecture. It provides a Master Routing Matrix that beautifully summarizes the myriad translocation signals used in both general cellular routing and specific Protein Sorting and Exocytosis. The table acts as an essential quick-reference guide, organizing complex biological data into clear, digestible categories. It meticulously maps out target destinations alongside their corresponding cellular pathways, specific signal types, and unique biochemical markers, providing a high-yield review tool.
The matrix clearly divides the Secretory and Cytoplasmic pathways. For the Secretory route, which dominates the mechanisms of Protein Sorting and Exocytosis, we see the familiar components. It outlines the N-terminal alpha-helix for initial ER entry, the C-terminal KDEL sequence for ER retention, and the unique carbohydrate Mannose 6-phosphate for lysosomal targeting. It also summarizes the apolar Stop-transfer sequences for membrane integration and structural 3D patches for secretory vesicles, encapsulating the diverse chemical vocabulary required for export.
Conversely, the bottom half of the matrix reviews the non-secretory signals. It reiterates the positively charged mitochondrial sequences, the internal basic KKKRK motif in the nucleus, and the C-terminal SKL signal in peroxisomes. Having all these markers in one matrix emphasizes the sheer volume of information a cell processes simultaneously. It underscores that while Protein Sorting and Exocytosis is a massive logistical operation, it exists alongside parallel, equally sophisticated routing systems that manage the cell’s internal organelles.
For college and medical students preparing for board exams, this matrix is an invaluable resource. The study of Protein Sorting and Exocytosis frequently requires memorizing these specific motifs. By understanding this summary table, students can quickly diagnose cellular misrouting in clinical vignettes. If a protein is found in the wrong compartment, consulting this Master Matrix of Protein Sorting and Exocytosis allows a student to easily identify which specific molecular signal or receptor has structurally failed, connecting basic biochemistry to clinical pathology.
Please read our Content Disclaimer Statement.
Check out our social media channels: