73. Membrane Transport Proteins: Structures and Mechanisms
A living cell is much like a biological fortress surrounded by an impenetrable lipid wall. Its daily survival strictly depends on highly regulated molecular gates to let essential nutrients inside and toxic waste out. This comprehensive structural deck directly illuminates the intricate architecture and vital functions of cellular transporters. The core purpose of this presentation is to detail the specific biochemical pathways and mechanical processes that govern cellular molecular uptake, providing an essential foundation for university and medical students studying modern cell biology.
Slide 1: Introduction to Membrane Transport Proteins
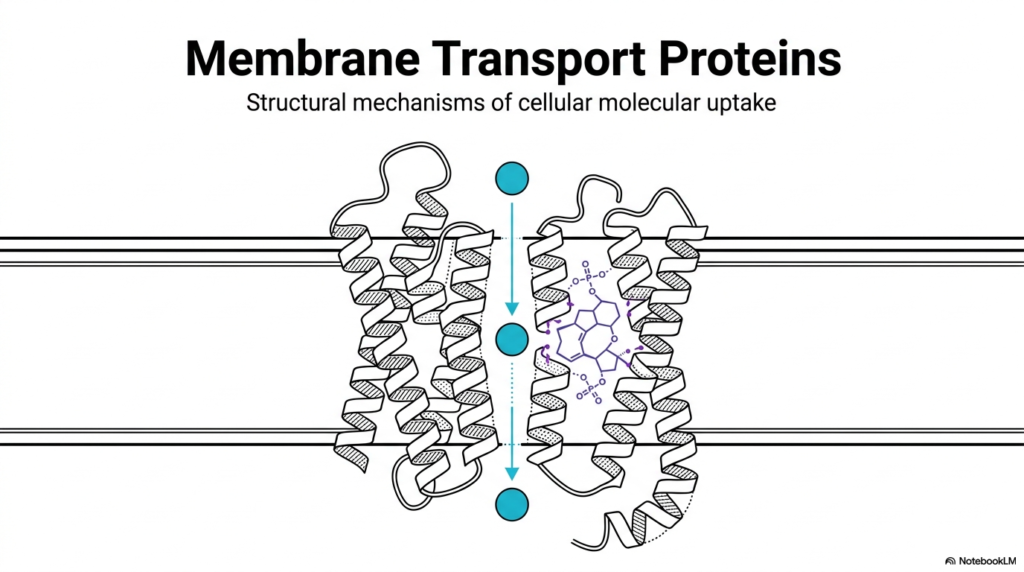
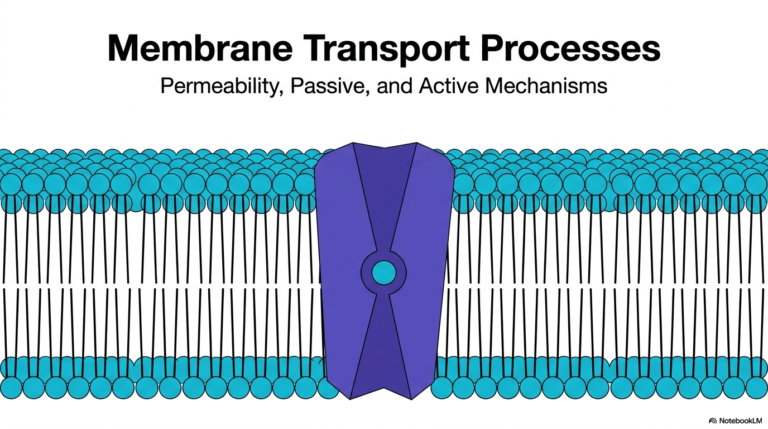
The foundational architecture of cellular life heavily relies on the intricate function of Membrane Transport Proteins. These complex molecular machines are embedded directly within the cell’s lipid bilayer, acting as highly selective gatekeepers. Without Membrane Transport Proteins, essential nutrients could not enter the intracellular space, and toxic metabolic byproducts would rapidly accumulate to lethal levels. The primary illustration highlights the elegant structural mechanisms that govern cellular molecular uptake. The visual reveals a precise cross-section of a biological membrane containing a complex transport channel.
By analyzing this specific structural conformation, observers can clearly see how alpha-helical protein structures form a dedicated pore through an otherwise impermeable lipid barrier. Membrane Transport Proteins typically organize into precise three-dimensional shapes that create a safe, hydrophilic pathway for specific molecules to traverse. In this structural overview, a targeted molecule passes safely through the hydrophobic core of the biological lipid bilayer. The surrounding protein structure physically shields the transiting molecule from the lipid environment, demonstrating the exact nature of these vital biochemical gateways.
Understanding these specific structures provides the critical basis for all subsequent biochemical topics related to cellular homeostasis and metabolism. Medical and university students must recognize that these macro-structures are never simply static tubes, but rather highly dynamic engines. They constantly respond to various chemical, physical, and electrical signals to maintain the delicate internal balance required for cellular survival. This opening visual perfectly sets the stage for exploring the diverse thermodynamic strategies employed across vastly different cellular environments and distinct tissue types.
As the foundational elements of mammalian cellular physiology, these biological gateways frequently represent primary targets for crucial pharmacological interventions. Countless modern medications function specifically by either inhibiting or enhancing the native activity of these channels. By mastering the fundamental architecture presented in this introductory model, aspiring scientists and medical professionals gain the essential context to understand complex metabolic diseases. This knowledge directly translates into a better comprehension of targeted drug therapies that artificially alter natural cellular molecular uptake.
Slide 2: Fundamental Mechanisms of Membrane Transport Proteins
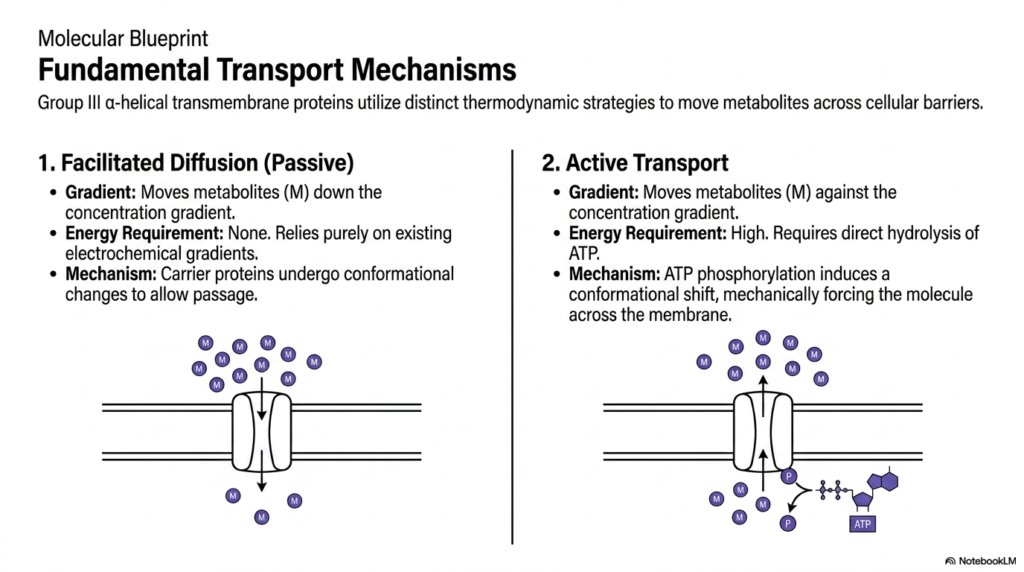
Cellular physiology relies on distinct thermodynamic strategies to transport metabolites across biological barriers, a process heavily mediated by Membrane Transport Proteins. The two primary mechanisms are passive facilitated diffusion and active transport. In facilitated diffusion, metabolites move strictly down their concentration gradient. This passive process requires zero external energy input, relying entirely on existing electrochemical gradients. The Membrane Transport Proteins involved act as specialized carrier molecules that undergo subtle conformational changes to allow passage through the lipid bilayer, ensuring rapid equilibration of vital metabolic substrates.
Conversely, active transport represents a biologically expensive but necessary strategy. This mechanism moves metabolites aggressively against their natural concentration gradient, forcing molecules from areas of low concentration to areas of high concentration. To achieve this demanding task, these specific Membrane Transport Proteins require a high-energy input, typically derived from the direct hydrolysis of cellular ATP. The chemical energy liberated from this potent reaction fuels the physical mechanical work required to move molecules against thermodynamic favorability.
The precise mechanism of active transport involves intricate biochemical steps. ATP phosphorylation directly induces a massive conformational shift within the protein structure. This structural alteration mechanically forces the target molecule across the cellular membrane. The distinction between passive and active mechanisms is absolutely paramount for students studying cellular bioenergetics. It highlights how cells strategically allocate their limited energy reserves only when necessary to maintain vital electrochemical gradients or concentrate essential metabolic substrates against opposing physical forces.
Visualizing these fundamental processes clarifies the mechanical reality of cellular metabolism. Group III alpha-helical transmembrane structures form the physical basis for both of these vital cellular pathways. Whether utilizing existing gradients or burning chemical energy to create new ones, the structural blueprint remains a marvel of evolutionary engineering. Grasping these distinct energetic requirements allows medical researchers to accurately predict how complex cellular systems will react under various conditions of metabolic stress, starvation, or acute energy depletion.
Slide 3: Secondary Active Transport and Membrane Transport Proteins
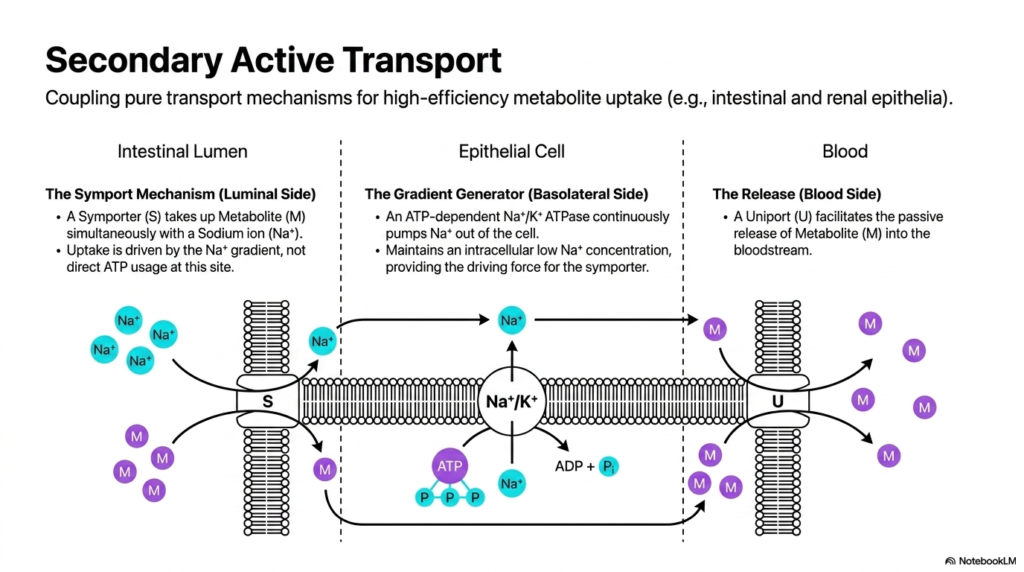
Secondary active transport demonstrates a brilliant cellular strategy for coupling pure transport mechanisms to achieve high-efficiency metabolite uptake. This process is particularly crucial in specialized tissues like the intestinal and renal epithelia. In these environments, distinct Membrane Transport Proteins work in a highly coordinated sequence. On the luminal side, a symport mechanism takes up a desired metabolite simultaneously with a sodium ion. Crucially, this specific uptake is driven entirely by the existing sodium gradient, not by direct ATP usage at the luminal site.
To maintain the driving force for this symporter, the cell relies on a separate gradient generator located on the basolateral side of the epithelial cell. Here, an ATP-dependent sodium-potassium ATPase continuously pumps sodium ions out of the intracellular space. This relentless pumping action maintains a consistently low intracellular sodium concentration. The Membrane Transport Proteins functioning as gradient generators consume significant ATP to ensure the luminal symporter always has the thermodynamic driving force required to bring important nutritional metabolites into the cell.
The final stage of this complex transepithelial journey occurs at the blood side of the cellular barrier. A specialized uniport mechanism facilitates the passive release of the concentrated metabolite directly into the bloodstream. This carefully choreographed dance between three separate Membrane Transport Proteins ensures unidirectional, highly efficient nutrient absorption. The system perfectly illustrates how cells can invest energy in one specific location to drive passive, gradient-dependent transport in another completely distinct region of the extensive cellular membrane.
By mastering this coupled transport sequence, medical students can better understand the exact physiological basis of oral rehydration therapies and dangerous nutrient malabsorption disorders. The precise spatial distribution of these distinct molecular machines across the apical and basolateral membranes is not random, but highly evolved. It creates a unidirectional flow of essential nutrients from the gut or kidney lumen directly into the systemic circulation, a process vital for overall organismal survival and daily metabolic maintenance across all mammalian species.
Slide 4: Isoforms of the GLUT Family Membrane Transport Proteins
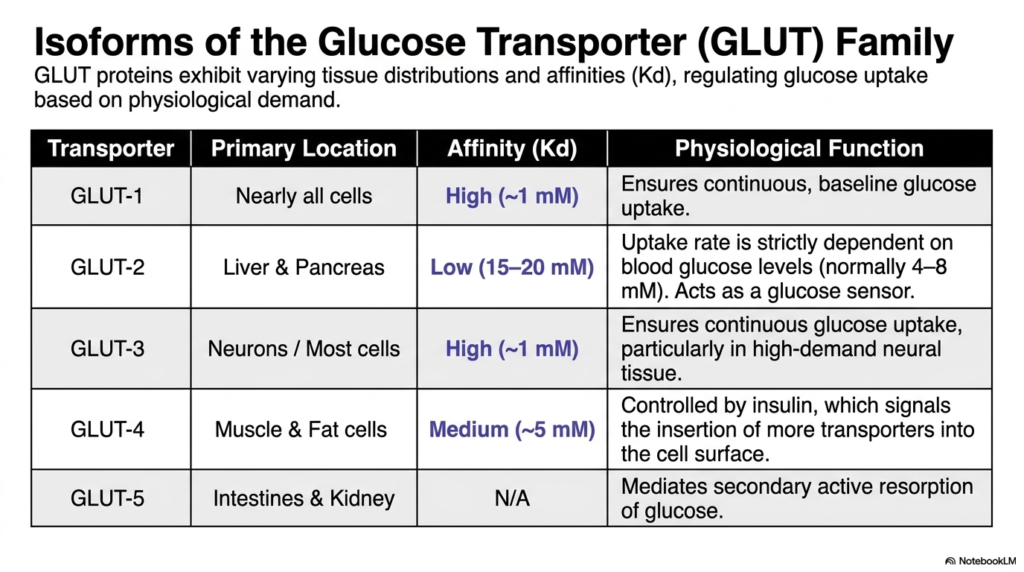
The complex regulation of systemic glucose uptake perfectly illustrates the functional diversity among Membrane Transport Proteins. The Glucose Transporter, or GLUT family, exhibits varying tissue distributions and distinct binding affinities to carefully regulate metabolic fuel based on specific physiological demands. Understanding these minor structural variations is essential for comprehending human metabolism and complex diseases like diabetes. Different tissues require vastly different baselines of glucose supply, and the evolutionary adaptation of these specific Membrane Transport Proteins reflects that biological reality perfectly.
GLUT-1 and GLUT-3 are both high-affinity transporters with a very low dissociation constant around one millimolar. GLUT-1 is located in nearly all mammalian cells, ensuring a continuous, reliable baseline of glucose uptake regardless of minor blood sugar fluctuations. Similarly, GLUT-3 is highly expressed in neurons and other critical tissues, ensuring continuous glucose uptake for high-demand neural processing. These specific Membrane Transport Proteins guarantee that the brain and other vital organs never face a critical shortage of their primary metabolic fuel.
In stark contrast, GLUT-2 serves an entirely different physiological function in the liver and pancreas. It functions as a low-affinity transporter, meaning its cellular uptake rate is strictly dependent on circulating blood glucose levels. This specific biochemical characteristic allows GLUT-2 to act as a highly effective physiological glucose sensor, triggering appropriate hormonal responses when blood sugar spikes. Meanwhile, GLUT-4 is a medium-affinity transporter located primarily in muscle and fat cells and is distinct in that its surface expression is directly controlled by insulin signaling.
Finally, GLUT-5 functions uniquely within the intestines and kidneys to mediate the secondary active resorption of specific biological sugars like fructose. By analyzing this comprehensive isoform table, students can easily appreciate how a single class of structural molecules diversifies to meet the highly complex needs of organisms across different organ systems. The careful distribution of high-, low-, and medium-affinity transporters ensures that circulating systemic glucose is optimally partitioned between immediate energetic demands in the central nervous system and long-term storage in muscular tissues.
Slide 5: Structural Deep Dive into GLUT-1 Membrane Transport Proteins
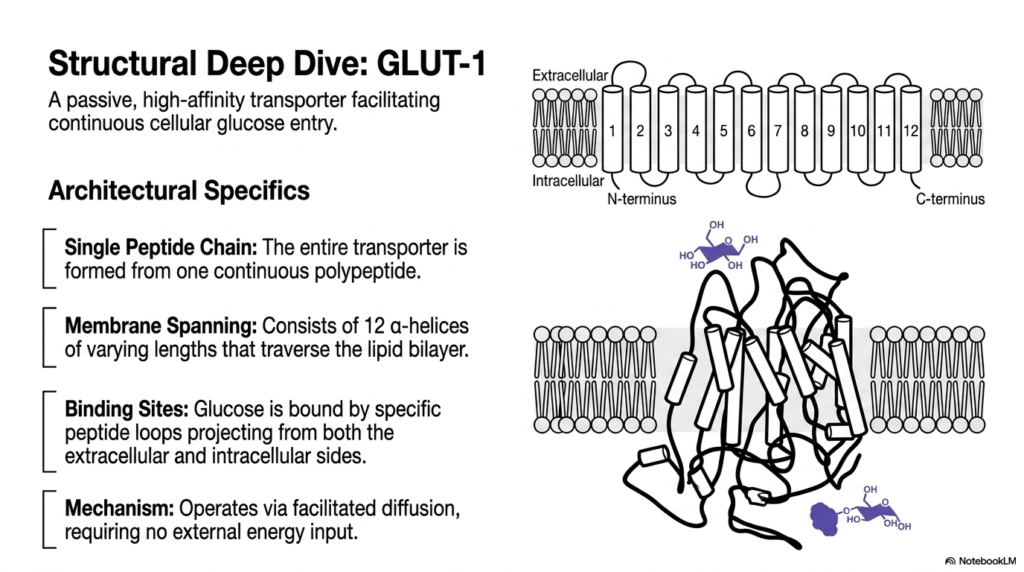
Taking a closer look at architectural specifics reveals the elegant biological design of the GLUT-1 transporter. This massive molecule is a classic example of a passive, high-affinity Membrane Transport Protein that facilitates continuous cellular glucose entry. The entire structural apparatus is formed from a single, continuous polypeptide chain. Despite being a single continuous peptide, it meticulously folds into a highly complex three-dimensional shape that is absolutely necessary for its critical biological function. This specific folding allows these specialized Membrane Transport Proteins to span the cellular barrier effectively.
The membrane-spanning region of GLUT-1 consists of exactly twelve distinct alpha-helices of varying lengths. These strong helical structures intricately traverse the hydrophobic lipid bilayer, forming a protective, hydrophilic internal pore. The metabolic substrate, glucose, is tightly bound by specific peptide loops projecting outward from both the extracellular and intracellular sides of the channel. These specialized binding sites provide the exceptional selectivity that defines these specialized Membrane Transport Proteins, ensuring only the correct metabolic carbohydrate gains entry into the cellular interior.
Mechanistically, GLUT-1 operates entirely via passive facilitated diffusion. This specific mechanism means the functional transporter requires absolutely no external energy input to operate. Instead, the inherent thermal energy of the biological system and the existing chemical gradient drive the movement of glucose molecules. When a target glucose molecule binds to the extracellular loops, it instantly induces a subtle but highly significant conformational shift in the twelve transmembrane helices. This physical rearrangement alternately exposes the central binding site to the cell’s interior and exterior.
For medical students analyzing biological structures, GLUT-1 is an ideal introductory model of carrier-mediated cellular transport. The provided visual rendering clearly demonstrates how the complex intertwining of twelve transmembrane alpha-helices creates a functional, highly selective gateway. This structural deep dive clearly clarifies how a seemingly simple linear peptide chain can spontaneously fold and evolve into a sophisticated, highly specific molecular machine that sustains mammalian cellular respiration by ensuring a steady, uninterrupted supply of essential carbohydrates to the active intracellular environment.
Slide 6: Aquaporin-1 and Highly Selective Membrane Transport Proteins
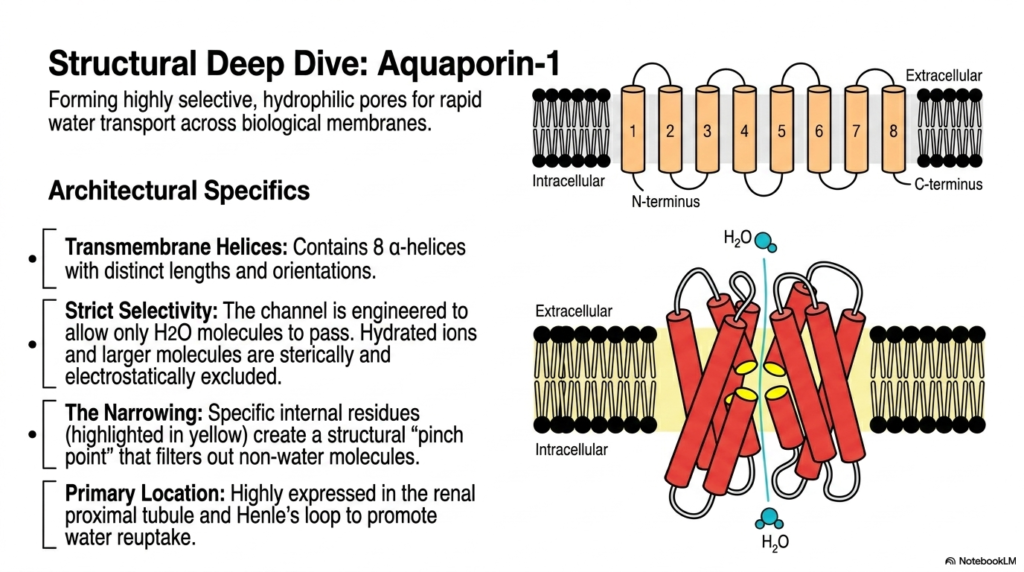
Water movement across biological barriers is a fundamental requirement for terrestrial life and is largely governed by specialized Membrane Transport Proteins known as aquaporins. Aquaporin-1 forms highly selective, hydrophilic pores designed specifically for the rapid, continuous transport of water across cellular membranes. Structurally, the precise architecture of Aquaporin-1 differs significantly from that of typical glucose transporters. This vital channel contains exactly eight transmembrane alpha-helices with distinct physical lengths and complex spatial orientations. These diverse Membrane Transport Proteins demonstrate how unique molecular folding patterns yield vastly different chemical selectivities.
The single most remarkable feature of Aquaporin-1 is its strict, uncompromising selectivity. The structural transmembrane channel is perfectly engineered to allow only a single water molecule to pass through in a precise single-file line. Hydrated cellular ions and larger biological molecules are strictly excluded through both mechanical steric and electrical electrostatic mechanisms. This phenomenal structural filtering ability makes these Membrane Transport Proteins incredibly efficient at regulating osmotic balance without inadvertently altering the critical electrochemical gradients of essential cellular ions such as sodium and potassium.
This exceptional molecular selectivity is achieved mechanically by a specific, physical narrowing deep within the internal pore structure. Specific internal amino acid residues act cooperatively to create a tight structural pinch point that physically and chemically filters out any non-water molecules. Water molecules must precisely orient their dipoles to pass through this narrow gateway, ensuring that even tiny protons cannot piggyback their way through the open channel. This ingenious mechanism prevents the deadly, rapid collapse of cellular proton gradients during massive water uptake events.
Physiologically, Aquaporin-1 is highly expressed in the renal proximal tubule and the descending limb of Henle’s loop. In these highly specific anatomical locations, the permanent baseline channels are absolutely necessary to promote continuous, massive reuptake of baseline water from the forming urine directly back into the systemic bloodstream. Understanding the distinct structural ingenuity of these ubiquitous channels provides medical and university biochemistry students with crucial insights into human osmoregulation, renal physiology, and the fundamental physical principles of strict molecular exclusion in living biological systems.
Slide 7: Endocrine Regulation of Aquaporin-2 Membrane Transport Proteins
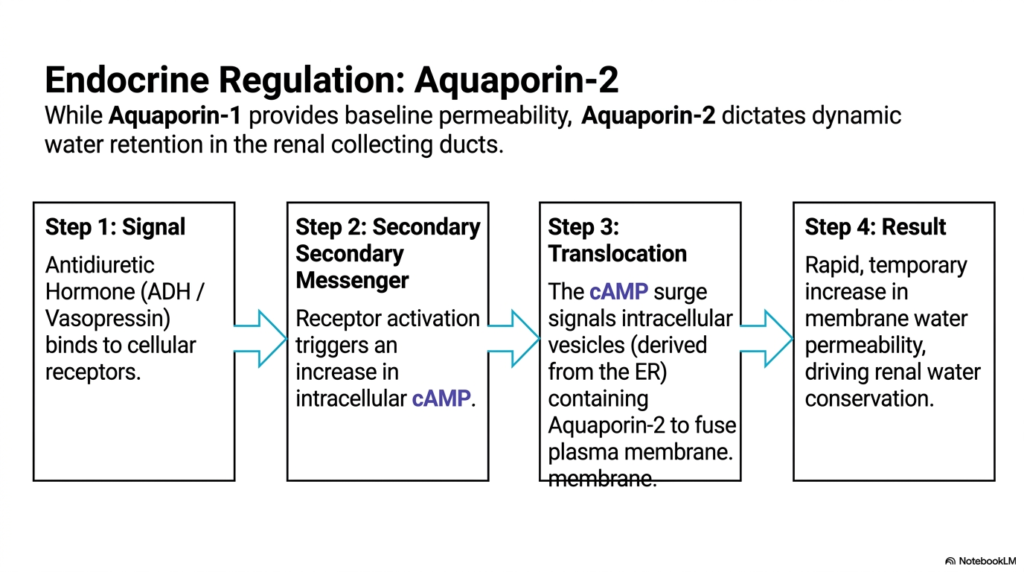
While some channels provide continuous baseline permeability, others are dynamically regulated by the endocrine system. The physiological regulation of Aquaporin-2 offers a brilliant demonstration of exactly how Membrane Transport Proteins are controlled by complex chemical signaling cascades. Unlike its unregulated counterpart, Aquaporin-2 dictates dynamic, temporary water retention specifically within the critical renal collecting ducts. This process ensures the body can rapidly adjust its fluid balance in response to systemic hydration levels. Such tightly regulated Membrane Transport Proteins actively prevent massive, fatal fluid loss during periods of severe dehydration.
The regulatory biochemical sequence initiates with a highly specific chemical signal. Antidiuretic Hormone, also widely known as Vasopressin, binds directly to specialized cellular receptors located strictly on the basolateral membrane of the kidney cells. This initial receptor activation instantly triggers a potent secondary messenger cascade, resulting in a marked increase in intracellular cyclic AMP. This rapid increase in cAMP acts as the crucial biological signal that actively alters the cellular localization of specific Membrane Transport Proteins stored safely within the intracellular environment.
The third operational step involves rapid, targeted protein translocation. The potent cAMP surge forcefully signals intracellular vesicles containing pre-formed Aquaporin-2 channels to immediately move toward and fuse directly with the apical plasma membrane. These specialized transport vesicles, originally derived from the endoplasmic reticulum, effectively deploy the critical water channels exactly where they are needed most. The mechanical membrane fusion seamlessly inserts the functional channels into the cellular barrier, instantly changing the physical properties of the renal collecting duct epithelial lining.
The final physiological result is a rapid, temporary, and highly significant increase in overall apical membrane water permeability. This complex chemical cascade powerfully drives vital renal water conservation, allowing the human body to highly concentrate urine and reabsorb precious fluid directly back into the systemic bloodstream. For advanced students in medical and biological disciplines, this dynamic sequence perfectly illustrates the seamless intersection of clinical endocrinology and fundamental cellular biochemistry. It definitively reveals how systemic hormonal signals directly manipulate the microscopic membrane architecture to achieve macroscopic physiological survival.
Slide 8: Classification of Active Membrane Transport Proteins and ATPases
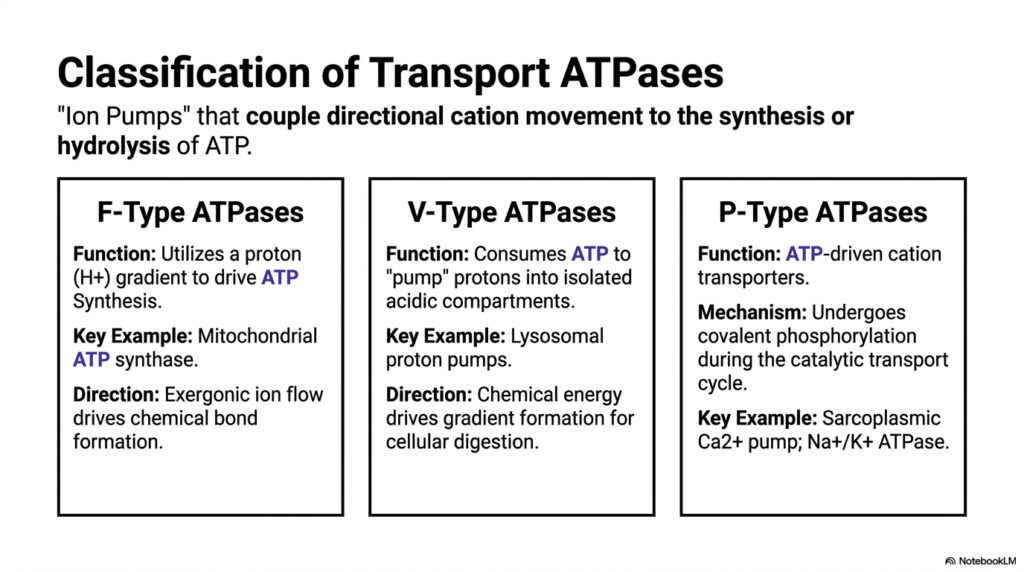
To comprehensively understand active biological transport, scholars must systematically classify the diverse enzymes that perform this thermodynamically demanding work. Transport ATPases consistently act as powerful biological ion pumps. These highly specialized Membrane Transport Proteins strictly couple the directional movement of cellular cations to the vital synthesis or direct hydrolysis of cellular ATP. This potent energetic coupling forms the absolute foundation of all cellular bioenergetics. By accurately categorizing these active Membrane Transport Proteins, students can better understand how different cellular organelles manage their vast energy reserves.
The first major structural classification is the F-Type ATPases. These remarkable molecular machines function primarily by using a strong biological proton gradient to directly drive ATP synthesis. The most famous physiological example is mitochondrial ATP synthase, which relentlessly powers almost all mammalian cellular activity. In this specific complex system, the purely exergonic flow of isolated ions down their gradient provides the mechanical rotational force necessary to drive chemical bond formation. These Membrane Transport Proteins essentially act as microscopic cellular turbines, spinning rapidly to generate vital energetic currency.
The second functional category comprises the V-Type ATPases, which generally run the typical F-Type mechanism in strict mechanical reverse. Instead of generating chemical energy, V-Type pumps rapidly consume cellular ATP to forcefully pump protons into highly isolated acidic compartments. A prime physiological example includes dynamic lysosomal proton pumps. By deliberately burning chemical energy, the active cell drives the formation of a massive proton gradient, creating the highly acidic, low-pH internal environment absolutely necessary for optimal cellular digestion and enzymatic waste degradation within specialized organelle compartments.
Finally, P-Type ATPases are classic ATP-driven cation transporters frequently found on the plasma membrane and the sarcoplasmic reticulum. Their defining mechanical feature is that they undergo direct, transient covalent phosphorylation during the active catalytic transport cycle. Key physiological examples include the Sarcoplasmic Calcium pump and the ubiquitous Sodium-Potassium ATPase. This comprehensive classification system enables biological scholars to accurately predict structural homology, underlying biochemical mechanisms, and the functional physiological consequences of various dynamic ion pumps across vastly different sectors of the complex cellular landscape.
Slide 9: Structural Architecture of P-Type Membrane Transport Proteins
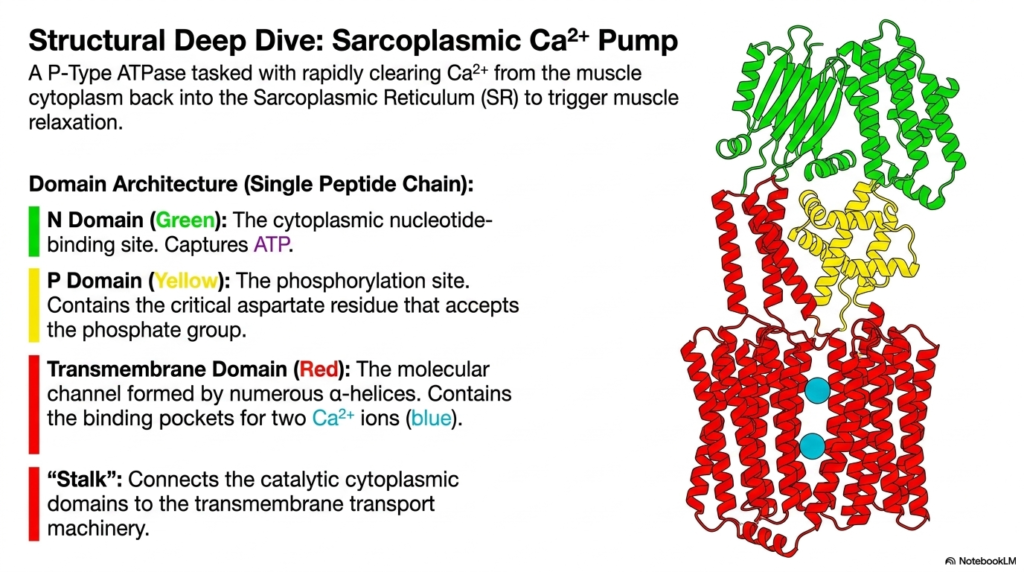
Examining the Sarcoplasmic Calcium Pump provides an incredible visual window into the highly complex domain architecture of classic P-Type enzymes. This specific, powerful pump rapidly clears toxic calcium ions from the active muscle cytoplasm and forcefully pushes them back into the Sarcoplasmic Reticulum. This rapid chemical clearance triggers essential, immediate muscle relaxation. The physical architecture of these specialized Membrane Transport Proteins perfectly reflects their intense energetic requirements. Like the glucose carrier, this massive molecular machine is built entirely from a single, highly complex peptide chain.
The massive domain architecture is beautifully segmented into specialized, highly distinct functional regions. The N Domain, clearly highlighted in green on the model, serves precisely as the cytoplasmic nucleotide-binding site. Its sole molecular purpose is to capture free cellular ATP from the surrounding cytoplasm. Adjacent to this structure is the P Domain, highlighted in yellow, which acts as the critical biological phosphorylation site. Highly evolved Membrane Transport Proteins strictly rely on this specific domain because it contains the absolutely essential aspartate residue that covalently accepts the terminal phosphate group during initial enzymatic activation.
Deep within the hydrophobic lipid bilayer lies the Transmembrane Domain, depicted vividly in red. This vital section constitutes the actual molecular transport channel, meticulously formed by numerous, tightly packed alpha-helices. It precisely contains the highly specific geometric binding pockets required to capture two distinct calcium ions directly from the cytoplasm. Connecting these functional active regions together is the stalk structure. This crucial structural component physically links the catalytic cytoplasmic domains to the complex transmembrane transport machinery, ensuring that the generated mechanical force properly translates into the opening of Membrane Transport Proteins.
By meticulously visualizing this distinct color-coded architecture, advanced students can clearly observe how specialized biochemical function is strictly partitioned within a single, very large macromolecule. The distinct structural separation between the energy-harvesting cytoplasmic domains and the actual physical transport channel is a universal hallmark of functional P-Type pumps. This structural blueprint beautifully clarifies how energy derived from ATP hydrolysis, located far above the lipid membrane, is mechanically transmitted downward to induce massive conformational changes that physically force calcium against its immense cellular concentration gradient.
Slide 10: The Catalytic Cycle of Active Membrane Transport Proteins
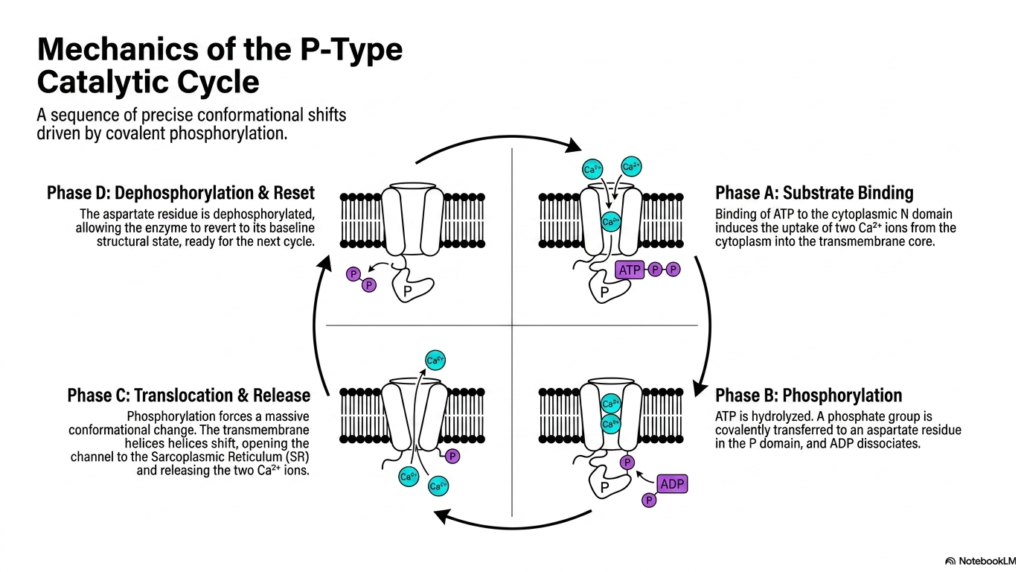
The true biochemical brilliance of P-Type ATPases lies precisely in their dynamic mechanical execution, beautifully illustrated by the mechanics of the active catalytic cycle. This sequence involves precise, massive conformational shifts driven entirely by rapid transient covalent phosphorylation. Understanding this specific continuous cycle reveals exactly how Membrane Transport Proteins accurately convert raw chemical energy into physical biological work. The cycle continuously oscillates through four distinct biochemical phases, ensuring rapid, unidirectional ion transport that resets itself perfectly for continuous, uninterrupted physiological operation within the highly active muscle cell.
The transport cycle inevitably begins with Phase A, which represents the initial substrate-binding event. During this specific phase, a single molecule of ATP strongly binds to the cytoplasmic N domain. Simultaneously, this specific structural conformation induces the rapid geometric uptake of two free calcium ions from the cytoplasm directly into the open transmembrane core. The full activation of these complex Membrane Transport Proteins strictly requires this precise, simultaneous binding of both the energetic chemical substrate and the physical target ions to successfully trigger the subsequent massive enzymatic reactions.
Phase B involves the highly critical structural phosphorylation step. The bound ATP substrate is rapidly hydrolyzed, and the severed terminal phosphate group is covalently transferred directly to the waiting aspartate residue in the adjacent P domain, causing the remaining ADP molecule to quickly dissociate. This covalent addition immediately forces Phase C, representing a massive structural translocation and cellular release event. The intense phosphorylation mechanically forces the bound transmembrane helices to violently shift, completely opening the deep channel to the Sarcoplasmic Reticulum. Consequently, the isolated Membrane Transport Proteins forcefully release the two bound calcium ions into the interior.
Finally, Phase D accurately represents the crucial enzyme dephosphorylation and structural reset mechanism. The critical phosphorylated aspartate residue is biochemically dephosphorylated, forcibly removing the bulky chemical tag from the domain. This vital molecular removal allows the entire enormous enzyme complex to naturally revert back to its baseline, low-energy geometric structural state. The cellular pump is now fully prepared and physically positioned for the very next cycle. Grasping this complex four-step oscillatory mechanism is absolutely paramount for medical students seeking to master the exact chemical foundations of skeletal muscle contraction and subsequent relaxation.
Please read our Content Disclaimer Statement.
Check out our social media channels: