58. Nucleotide Biosynthesis: A Biochemical Overview
Every living cell must constantly manufacture the very blueprint of its existence to survive and divide. This fundamental truth makes the study of metabolic pathways essential for future medical professionals and scientists. The core purpose of this slide deck is to break down complex biochemical networks into digestible concepts. It provides a comprehensive visual and textual guide to the synthesis, regulation, and clinical implications of DNA and RNA precursors.
Slide 1: Introduction to Nucleotide Biosynthesis
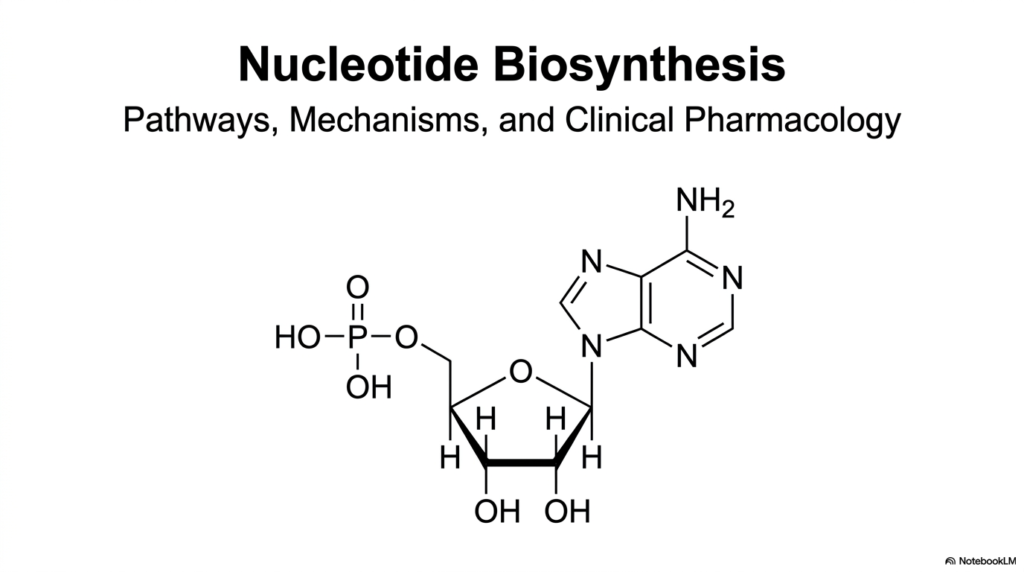
Welcome to the biochemical foundation of cellular replication and energy transfer. Nucleotide biosynthesis represents one of the most critical metabolic pathways across all forms of life. Without this essential machinery, cells simply cannot produce the DNA required for genetic inheritance, the RNA needed for protein translation, or the critical energy carriers like ATP. This initial presentation slide sets the stage for a highly detailed exploration into the intricate pathways, enzymatic mechanisms, and clinical pharmacology that define this complex field of study.
The image prominently features a classical nucleotide molecule, elegantly displaying its three foundational components. One can observe the central five-carbon sugar backbone, the energy-rich phosphate group, and the complex nitrogen-containing ring structure known as the nitrogenous base. Recognizing how these distinct chemical components connect is an absolute prerequisite for students. It serves as the baseline for mastering the sophisticated enzymatic reactions that dictate nucleotide biosynthesis.
Furthermore, the study of nucleotide biosynthesis is not just an academic exercise in memorizing chemical structures. It forms the biological basis for understanding how human diseases develop and how pharmacological interventions work. From targeting rapid cancer cell proliferation to managing painful metabolic disorders, the medical applications are vast. Medical and college students must grasp these baseline molecular shapes to appreciate how drugs bind to enzymes or interrupt metabolic flow.
To fully appreciate the scope of nucleotide biosynthesis, one must recognize its dual role in both cellular architecture and signaling. Nucleotides act as crucial coenzymes and signaling molecules that regulate countless physiological responses. Thus, this introductory slide does more than just show a static molecule; it introduces the very currency of biological energy and information. As the presentation progresses, the significance of this humble chemical structure will become incredibly clear.
Slide 2: The Core Pathways of Nucleotide Biosynthesis
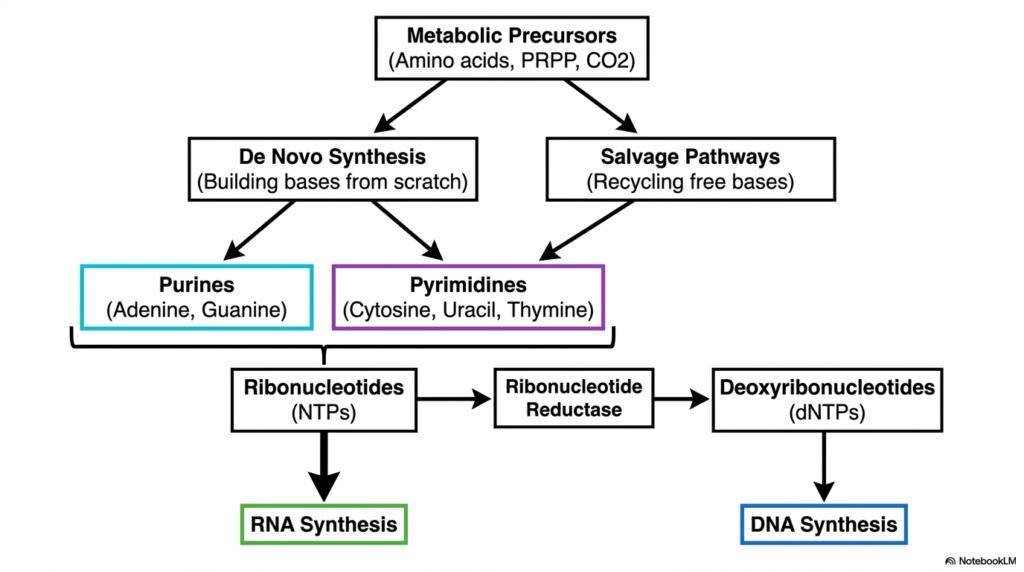
Understanding how cells acquire their genetic building blocks requires a big-picture view of the metabolic landscape. The entire framework of nucleotide biosynthesis can be conceptually divided into two major strategies: de novo synthesis and salvage pathways. De novo synthesis involves constructing purine and pyrimidine bases from scratch using basic metabolic precursors like amino acids, activated sugars, and carbon dioxide. This process is highly energy-intensive but absolutely vital for dividing cells.
Alternatively, the salvage pathways offer a metabolic shortcut. Instead of building molecules from basic raw materials, the cell aggressively recycles free bases released during cellular turnover. This biological recycling program significantly reduces the energy burden on the cell. The flowchart illustrates how both the de novo and salvage routes ultimately converge to produce ribonucleotides, the essential precursors for RNA synthesis.
Another crucial milestone in nucleotide biosynthesis is the conversion of ribonucleotides into deoxyribonucleotides. This highly regulated transformation is catalyzed by a remarkable enzyme known as ribonucleotide reductase. By stripping an oxygen atom from the ribose sugar, this enzyme provides the direct chemical precursors required for DNA synthesis. The clear visual separation between RNA and DNA precursor generation helps students map out the exact sequence of metabolic events.
This dual-pathway system ensures that cells maintain a constant and balanced pool of genetic material under varying physiological conditions. Whether a cell is rapidly dividing and relies heavily on constructing new bases, or it is resting and simply recycling old material, nucleotide biosynthesis adapts accordingly. Grasping this macroscopic flowchart is essential before diving into the specific enzymatic steps of purine and pyrimidine construction.
By organizing nucleotide biosynthesis into these distinct logistical routes, students can better anticipate how specific metabolic blocks will impact cellular function. For instance, understanding where ribonucleotide reductase operates provides immediate insight into how certain chemotherapeutic drugs halt DNA replication without directly destroying RNA production. This macroscopic perspective is the ultimate roadmap for navigating complex biochemical networks.
Slide 3: De Novo Purine Nucleotide Biosynthesis
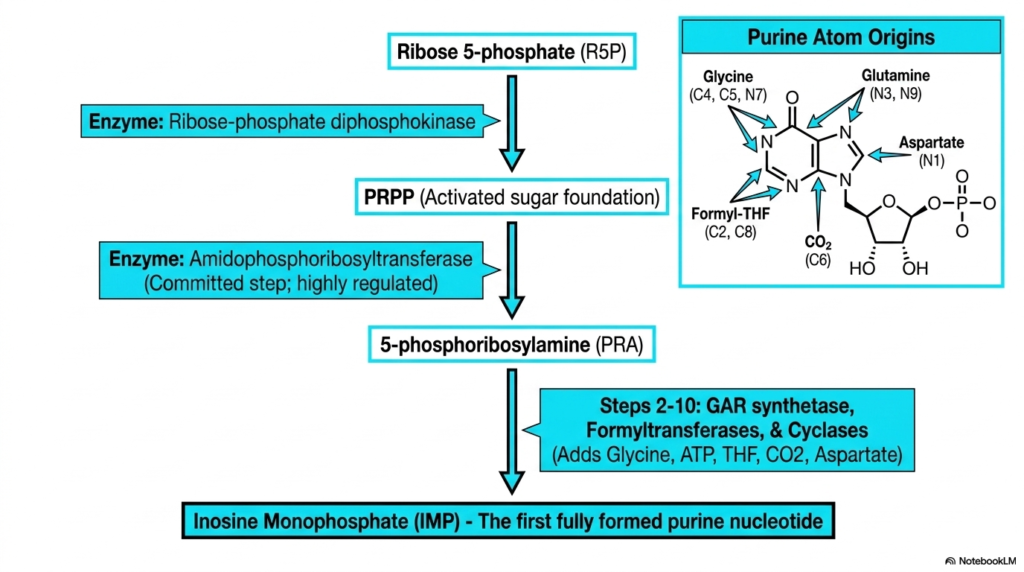
Diving into the specific mechanisms of nucleotide biosynthesis, the construction of purines requires an intricate, atom-by-atom assembly process. The de novo purine pathway is unique because the complex double-ring structure is built directly onto an activated sugar foundation, rather than being synthesized separately. This process begins with ribose 5-phosphate, which is converted into the crucial activated sugar foundation known as PRPP.
The committed, highly regulated step in this branch of nucleotide biosynthesis is catalyzed by the enzyme amidophosphoribosyltransferase. This enzyme facilitates the formation of 5-phosphoribosylamine, marking the point of no return for purine construction. From there, a sequence of multi-step enzymatic reactions systematically adds atoms to build the purine ring. The slide features a helpful diagram detailing the exact molecular origins of every atom in the purine framework.
Medical students must pay close attention to these atom sources during nucleotide biosynthesis. The purine ring incorporates nitrogen and carbon atoms donated by the amino acids glycine, glutamine, and aspartate. Additionally, carbon dioxide provides a crucial carbon atom, while formyl-tetrahydrofolate contributes two essential carbons. This heavy reliance on folate derivatives explains why folate deficiency so profoundly disrupts cellular division and replication.
The ultimate culmination of these complex assembly steps is the production of Inosine Monophosphate. As the first fully formed purine nucleotide, Inosine Monophosphate serves as the critical metabolic branch point. From this central hub, the cell will subsequently synthesize the specific adenine and guanine nucleotides required for life. This slide captures the systematic nature of biological ring assembly.
Mastering the intricacies of de novo purine construction is a rite of passage in biochemistry. The diagram highlights how multiple different metabolic pathways must intersect and cooperate to fuel nucleotide biosynthesis. If the cell lacks sufficient amino acids or active vitamin derivatives, the entire assembly line grinds to a halt. Recognizing these nutritional and enzymatic dependencies is paramount for understanding systemic metabolic diseases.
Slide 4: Branching Pathways in Nucleotide Biosynthesis
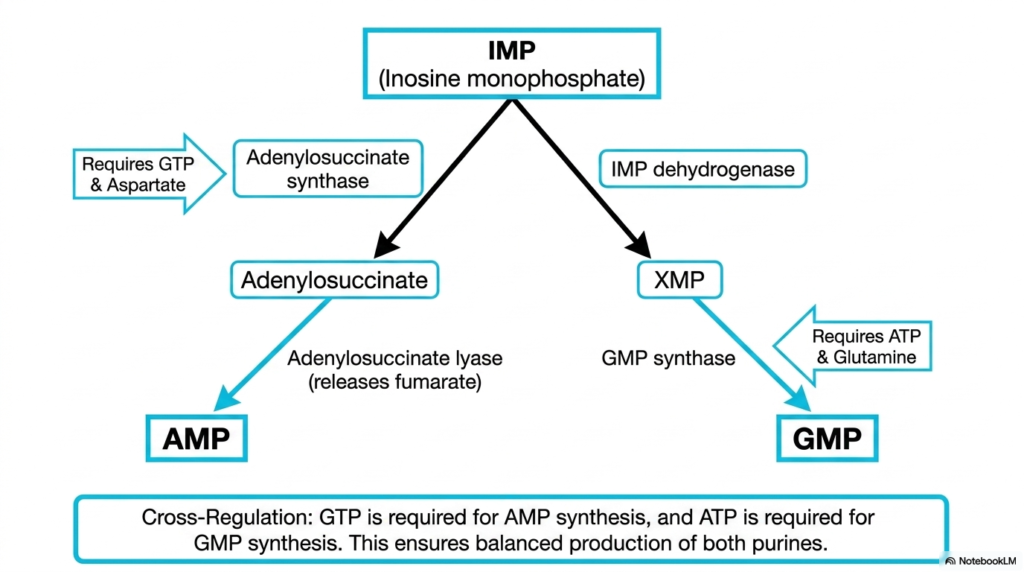
Once Inosine Monophosphate is successfully formed, the pathway of nucleotide biosynthesis faces a critical juncture. The cell must divide this precursor to generate balanced amounts of both Adenosine Monophosphate and Guanosine Monophosphate. This splitting mechanism ensures the genetic alphabet remains evenly stocked. The slide illustrates this bifurcation, highlighting the specific enzymatic steps and energy requirements for each respective branch.
To synthesize Adenosine Monophosphate, the pathway utilizes adenylosuccinate synthase and requires energy donated specifically by Guanosine Triphosphate. The amino acid aspartate also enters the reaction to donate a crucial amino group. Conversely, the route to Guanosine Monophosphate relies on Inosine Monophosphate dehydrogenase and GMP synthase. This specific pathway demands energy supplied by Adenosine Triphosphate and utilizes glutamine as its nitrogen donor.
This alternating energy requirement is a masterful stroke of biochemical regulation within nucleotide biosynthesis. Because the synthesis of one purine requires the fully energetic triphosphate form of the other, the cell inherently prevents the overproduction of either molecule. This elegant cross-regulation guarantees a balanced pool of purines, which is an absolute necessity for preventing catastrophic errors during DNA replication.
Understanding this reciprocal relationship is a cornerstone of mastering nucleotide biosynthesis. If a metabolic defect causes a massive spike in ATP, that energy will automatically drive the production of more GMP, bringing the system back into equilibrium. For students learning pharmacology, this branch point is also highly relevant, as enzymes like IMP dehydrogenase are prime targets for immunosuppressive medications designed to halt rapid cell division.
This precise balancing act reflects the high efficiency of cellular metabolism. The cell avoids wasting precious resources by strictly linking the production lines of its most important genetic building blocks. Students should take particular note of the enzymes operating at these branch points, as they represent vital control valves that govern the overall speed and direction of purine production.
Slide 5: Purine Catabolism and Nucleotide Biosynthesis
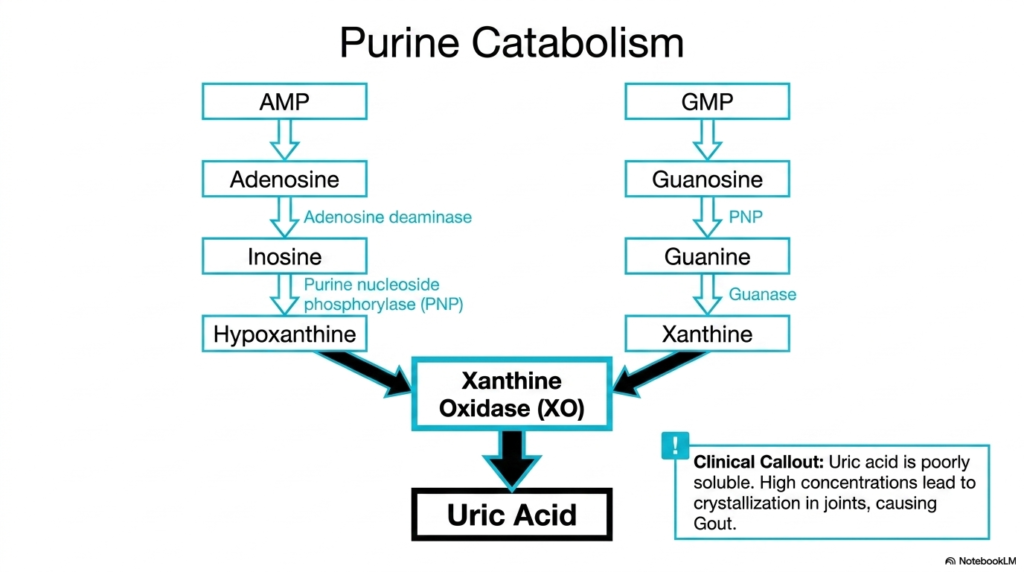
While the creation of metabolic building blocks is crucial, the breakdown process is equally significant. Although not directly a constructive phase of nucleotide biosynthesis, purine catabolism is the necessary biological counterbalance. When purines like AMP and GMP are degraded, they follow a highly specific dismantling pathway. The slide illustrates how both molecules are systematically stripped of their sugars and phosphate groups to yield free nitrogenous bases.
Adenosine Monophosphate is converted to inosine through the action of adenosine deaminase, and eventually becomes hypoxanthine. Guanosine Monophosphate is similarly processed down into the base guanine. Both of these intermediate bases converge on a common metabolic fate. The critical enzyme xanthine oxidase transforms these breakdown products first into xanthine, and finally into uric acid.
Unlike the highly soluble end products of other metabolic pathways, the uric acid produced here presents a unique biological challenge. Understanding this breakdown route is just as essential as understanding nucleotide biosynthesis because of its direct clinical implications. Uric acid is notoriously poorly soluble in human blood. When nucleotide degradation exceeds the body’s excretory capacity, uric acid concentrations spike dangerously high.
The clinical callout on the slide highlights the painful consequence of this metabolic imbalance: gout. The excess uric acid precipitates out of the blood, forming sharp, needle-like crystals in the joints. This inflammatory condition demonstrates why medical professionals must thoroughly study nucleotide biosynthesis and catabolism. By mapping out these degradative enzymes, students can easily identify where pharmacological inhibitors might be deployed to prevent uric acid accumulation.
The study of purine catabolism complements the broader topic of genetic precursor management. A healthy cell must constantly balance the rapid assembly of new molecules with the safe disposal of the old. This equilibrium ensures that the cellular environment remains free of toxic precipitates while maintaining a steady flux of metabolic intermediates.
Slide 6: The Salvage Pathway in Nucleotide Biosynthesis
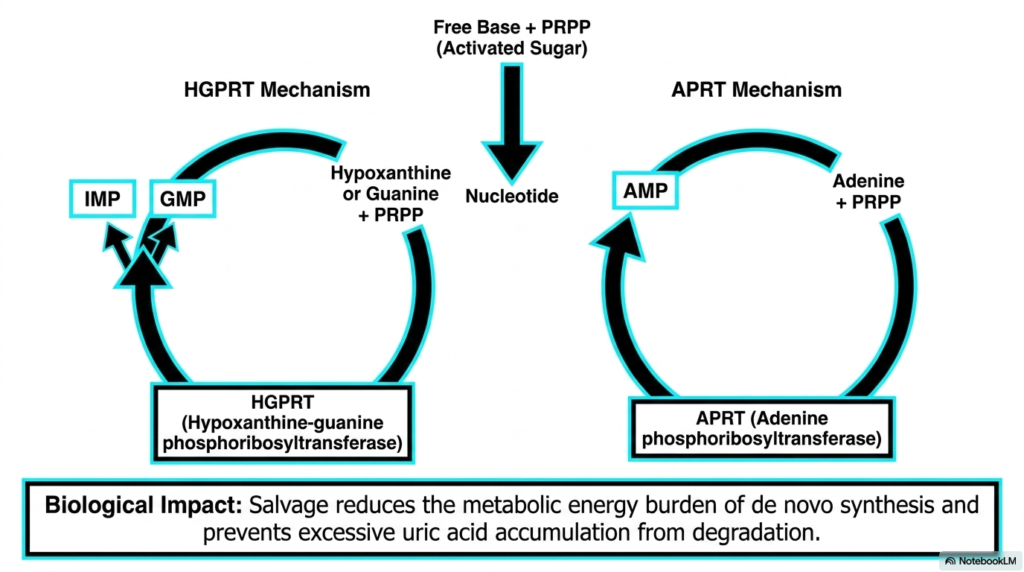
Nature is highly efficient, and the salvage pathway is a prime example of biological recycling. Within the grand scheme of nucleotide biosynthesis, constructing purines from scratch requires massive amounts of cellular energy. To minimize this metabolic burden, cells deploy highly specialized enzymes to rescue free bases before they are degraded into uric acid. This presentation slide details the two primary mechanisms responsible for this essential chemical rescue mission.
The first critical enzyme highlighted is Hypoxanthine-guanine phosphoribosyltransferase, commonly abbreviated as HGPRT. This remarkable protein intercepts free hypoxanthine or guanine bases and forcefully reattaches them to an activated sugar molecule. This direct chemical fusion immediately regenerates fully functional Inosine Monophosphate or Guanosine Monophosphate. The parallel enzyme, Adenine phosphoribosyltransferase, performs the exact same rescuing operation for free adenine molecules.
The biological impact of these salvage mechanisms within nucleotide biosynthesis cannot be overstated. By heavily relying on recycling, the cell drastically reduces the immense energy consumption otherwise demanded by de novo synthesis. Furthermore, by pulling free bases back into the functional genetic pool, the salvage pathway actively prevents the dangerous over-accumulation of uric acid that leads to severe metabolic disorders.
This slide underscores the interconnected nature of cellular economy and nucleotide biosynthesis. A failure in these specific recycling enzymes does not merely cause an energy shortage; it triggers catastrophic systemic disease due to unregulated waste buildup. Medical students must recognize these salvage enzymes not just as biochemical trivia, but as critical guardians of metabolic stability that protect the brain and joints from toxic damage.
Ultimately, the cellular preference for salvage over de novo construction dictates much of our baseline metabolic activity. Certain tissues, particularly within the central nervous system, rely almost exclusively on this recycling pathway to maintain their genetic material. This specific tissue dependency explains why inherited defects in these recycling enzymes present with such severe and targeted neurological symptoms.
Slide 7: Pyrimidine De Novo Nucleotide Biosynthesis
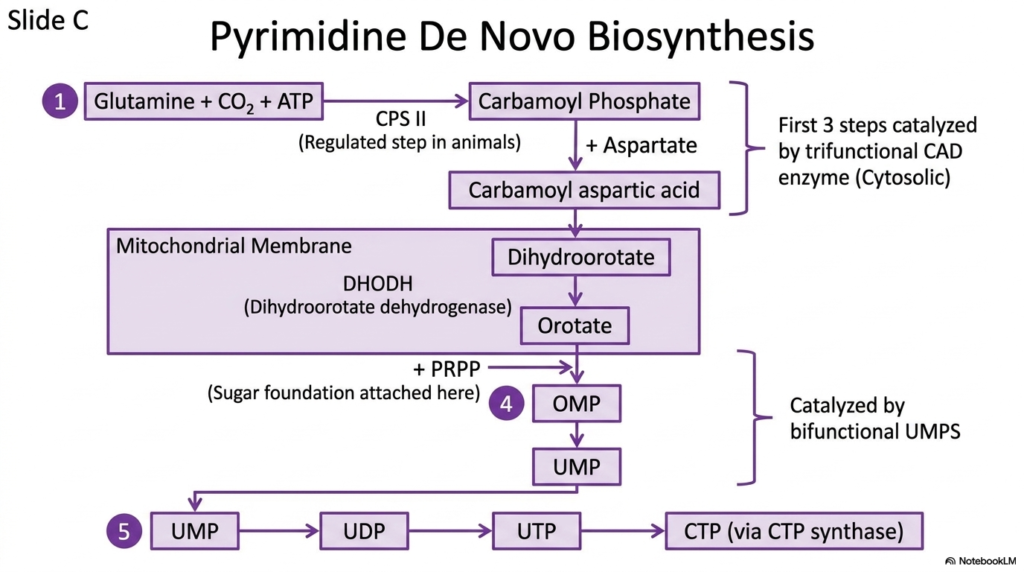
Shifting focus to the other half of the genetic alphabet, this slide details the assembly of pyrimidines. The strategy for pyrimidine nucleotide biosynthesis differs radically from purine construction. Instead of building the ring directly onto a sugar molecule, the cell synthesizes the entire pyrimidine ring independently first. Only after the ring is completely formed does the cell attach it to the activated sugar foundation.
The pathway begins with glutamine, carbon dioxide, and ATP combining to form carbamoyl phosphate. This regulated initial step is driven by the enzyme Carbamoyl phosphate synthetase II. The slide clearly highlights that the first three steps of this process are managed by a single, massive trifunctional enzyme known as CAD. This physical grouping of enzymatic activities dramatically accelerates the early stages of nucleotide biosynthesis.
Following these cytosolic reactions, the intermediate molecule travels to the mitochondrial membrane to interact with dihydroorotate dehydrogenase. This is the only step in pyrimidine nucleotide biosynthesis that does not occur in the cytosol. The resulting orotate molecule is finally linked to the sugar foundation to create Orotidine Monophosphate. This crucial connection is catalyzed by another multifunctional powerhouse, the bifunctional UMPS enzyme.
The pathway concludes with the generation of Uridine Monophosphate. From this foundational pyrimidine, the cell sequentially phosphorylates the molecule to build UDP and UTP. Finally, UTP can be transformed into CTP via the action of CTP synthase. This step-by-step schematic provides a clear roadmap of pyrimidine construction, highlighting the unique compartmentalization and multifunctional proteins involved.
Grasping the structural differences in how purines and pyrimidines are formed is essential for mastering metabolic biochemistry. The unique base-first approach of the pyrimidine pathway presents an entirely different set of regulatory checkpoints and enzymatic targets. For students, comparing these contrasting assembly strategies solidifies their overarching understanding of cellular metabolism.
Slide 8: Pyrimidine Catabolism and Nucleotide Biosynthesis
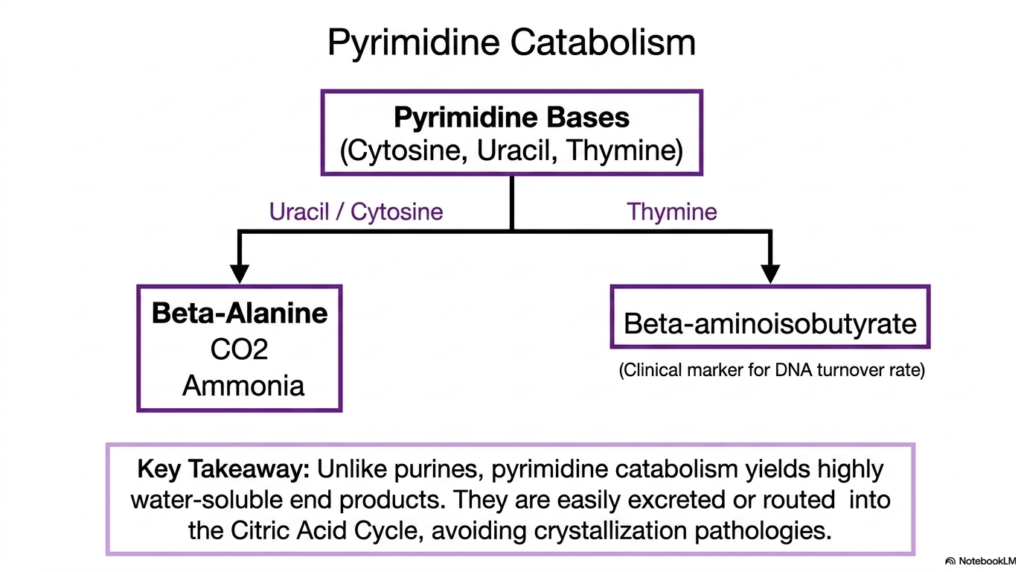
Just as purines require a dedicated disposal system, the breakdown of pyrimidines is a vital component of cellular maintenance. While nucleotide biosynthesis focuses on creating the complex rings required for life, pyrimidine catabolism safely deconstructs them. This slide illustrates the relatively straightforward degradation routes for the three main pyrimidine bases: cytosine, uracil, and thymine. The metabolic destiny of these molecules stands in stark contrast to their purine counterparts.
When the cell dismantles uracil and cytosine, the enzymatic processing yields beta-alanine, carbon dioxide, and ammonia. Conversely, the breakdown of thymine generates a specific molecule called beta-aminoisobutyrate. Medical professionals often utilize beta-aminoisobutyrate as a highly reliable clinical marker to measure the rate of DNA turnover in a patient. Monitoring this byproduct allows clinicians to assess how rapidly cells are dying and degrading their genetic material.
The key takeaway highlighted on this slide showcases a crucial metabolic advantage. Unlike the breakdown of purines, which generates dangerous and poorly soluble uric acid, pyrimidine catabolism yields highly water-soluble end products. These simple breakdown molecules are easily excreted by the kidneys without any risk of crystallization. Alternatively, the cell can seamlessly route them directly into the Citric Acid Cycle for energy production.
Understanding this efficient degradative process provides a more complete picture of nucleotide biosynthesis. Because the cell does not face severe toxic consequences when destroying pyrimidines, the regulatory pressures governing their disposal are entirely different. Students must appreciate this fundamental biochemical difference, as it explains why there are virtually no common inflammatory diseases associated with pyrimidine degradation.
The elegant simplicity of this catabolic pathway highlights the high adaptability of human metabolism. By converting complex genetic structures into simple energy intermediates, the cell prevents waste buildup and maximizes its energetic efficiency. This functional recycling ensures that the massive energy investment required by nucleotide biosynthesis is partially recovered during their eventual destruction.
Slide 9: Comparing Strategies in Nucleotide Biosynthesis
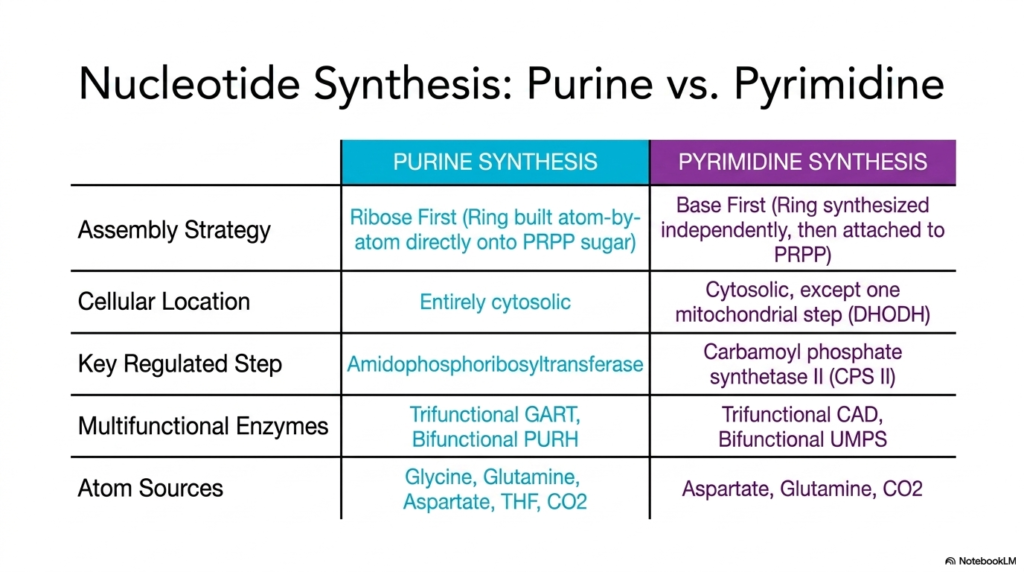
To truly master metabolic networks, one must be able to contrast competing biochemical pathways. This effective summary table directly compares the defining characteristics of purine and pyrimidine nucleotide biosynthesis. By placing these pathways side-by-side, the slide highlights the distinct evolutionary and structural strategies the cell employs to construct its genetic material.
The most profound difference lies in the fundamental assembly strategy. Purine nucleotide biosynthesis utilizes a ribose-first approach, meticulously building the complex double ring atom-by-atom directly onto the activated sugar. In stark contrast, pyrimidines use a base-first method, synthesizing the entire single ring independently before finally attaching it to the sugar foundation. This core mechanistic difference dictates the entirely different sets of enzymes required for each pathway.
The table also outlines the varying cellular locations and regulatory checkpoints. While purine synthesis is entirely cytosolic, pyrimidine construction features one unique mitochondrial step. Furthermore, the rate-limiting, highly regulated enzymes differ entirely. Purines rely on amidophosphoribosyltransferase, while pyrimidines are governed by Carbamoyl phosphate synthetase II. Both pathways heavily utilize massive multifunctional enzymes to streamline their complex chemical reactions.
Finally, the comparison of atom sources provides crucial insight into the nutritional requirements of nucleotide biosynthesis. While both pathways require the amino acid glutamine, purines uniquely demand vast quantities of glycine and folate derivatives. This categorical breakdown is an invaluable study tool for medical students. It transforms dense metabolic details into an easily digestible, comparative framework perfect for exam preparation.
By analyzing these pathways simultaneously, the underlying logic of cellular metabolism becomes apparent. The cell intentionally utilizes entirely separate regulatory mechanisms and molecular building blocks to prevent catastrophic metabolic traffic jams. This elegant separation ensures that the synthesis of one base type does not inadvertently deplete the raw materials needed for the other, guaranteeing genetic stability.
Slide 10: The Redox Chain of Nucleotide Biosynthesis
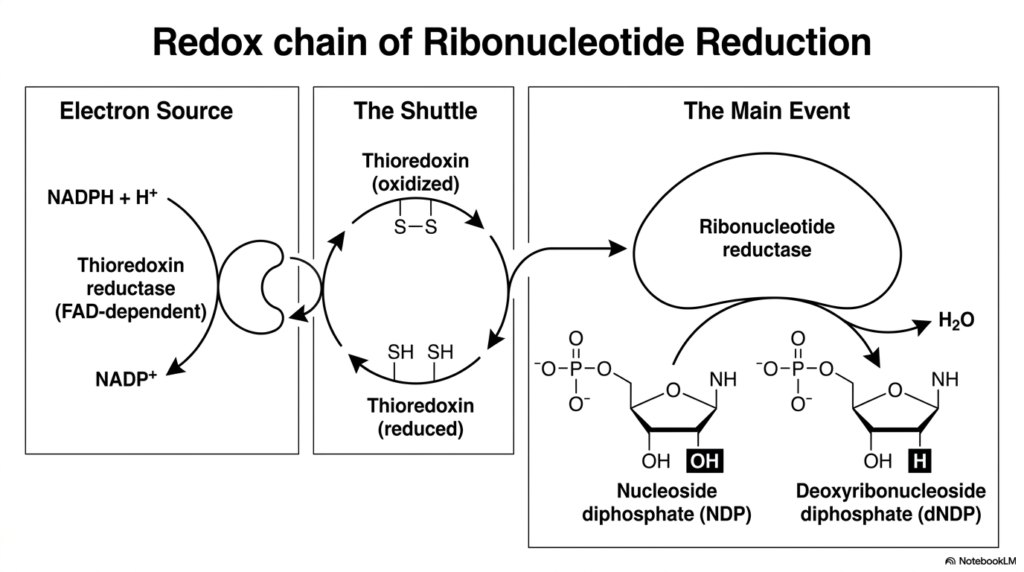
The transition from RNA precursors to DNA precursors represents one of the most chemically fascinating events in biology. This slide illustrates the complex redox chain that drives this specific phase of nucleotide biosynthesis. To build DNA, the cell must physically remove a single oxygen atom from a ribonucleotide. This incredibly difficult chemical feat is achieved through a multi-stage electron transfer process powered by the master enzyme, ribonucleotide reductase.
The process relies on a continuous flow of electrons, beginning with the ultimate electron source: NADPH. The enzyme thioredoxin reductase utilizes these electrons to convert oxidized thioredoxin into its active, reduced state. This reduced thioredoxin acts as a highly mobile biological shuttle. It carries the vital electrons directly to the main event, delivering them straight into the active site of ribonucleotide reductase.
Once ribonucleotide reductase is armed with these electrons, it can perform the critical step of nucleotide biosynthesis. It attacks the nucleoside diphosphate, cleaving the carbon-oxygen bond on the sugar ring. Water is released as a byproduct, and a deoxyribonucleoside diphosphate is born. This tightly coordinated bucket-brigade of electrons is the sole biological mechanism for generating the raw materials needed for DNA replication.
Without this elegant redox chain, cellular division would be physically impossible. The slide breaks down this complex electron transport system into three distinct, easy-to-understand phases: the source, the shuttle, and the main event. For biochemistry students, visualizing this exact flow of electrons demystifies how cells magically transform fragile RNA components into the highly stable building blocks of DNA.
Understanding this precise electron flow is also critical for modern pharmacology. Because this redox chain is the absolute bottleneck for nucleotide biosynthesis, interrupting any stage of this process immediately halts cellular proliferation. Researchers continuously study the intricate interactions between these specific redox proteins to develop more effective chemotherapies that can strategically starve cancer cells of their genetic material.
Slide 11: The Mechanism of Ribonucleotide Reductase in Nucleotide Biosynthesis
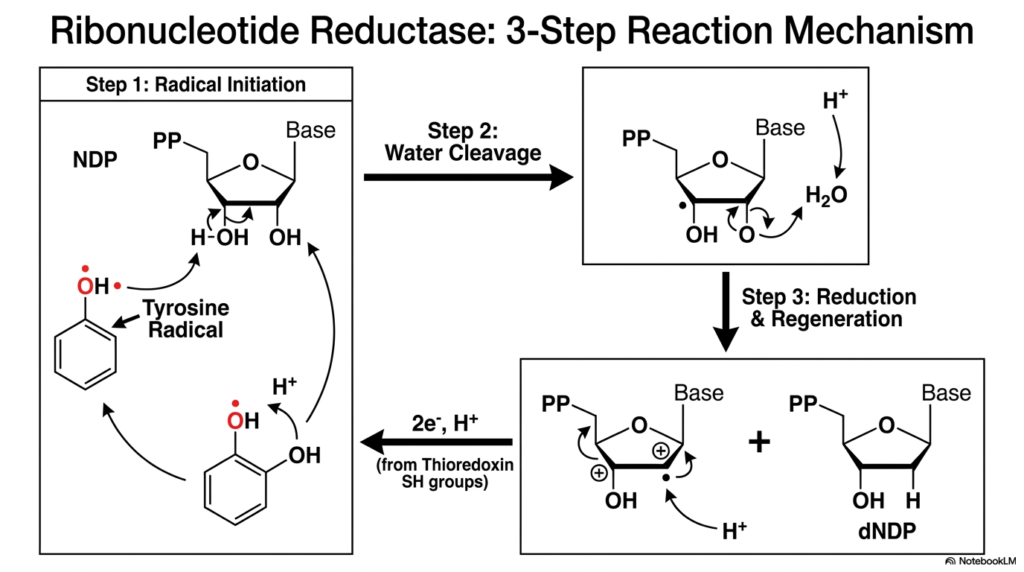
Diving deeper into the chemistry of DNA precursor generation, this slide visualizes the exact, atomic-level mechanism of ribonucleotide reductase. This enzyme executes the most chemically demanding reaction in all of nucleotide biosynthesis. Removing an oxygen atom from a highly stable ribose ring requires the dangerous but necessary use of free radicals. The presentation slide breaks this violent chemical reaction down into three distinct, sequential steps.
The reaction begins with Step 1: Radical Initiation. The enzyme utilizes a specialized tyrosine amino acid that harbors a highly reactive, unpaired electron. This tyrosine radical aggressively attacks the sugar ring of the nucleotide, abstracting a hydrogen atom and transferring the radical state directly onto the substrate. This bold chemical maneuver destabilizes the ring, setting the stage for the crucial oxygen removal required in nucleotide biosynthesis.
Step 2 features the actual Water Cleavage. With the substrate now highly unstable, the enzyme orchestrates the release of the targeted hydroxyl group. The oxygen atom combines with available protons to form a molecule of water, which is then ejected from the active site. Following this cleavage, Step 3 involves the final Reduction and Regeneration. Electrons delivered by the thioredoxin shuttle pour into the active site to resolve the radical state.
The fully formed deoxyribonucleotide is released, and the crucial tyrosine radical is regenerated, ready to attack the next molecule. This step-by-step schematic is vital for advanced medical and biochemistry students studying nucleotide biosynthesis. It vividly illustrates how enzymes can safely harness and control dangerous free radical chemistry to perform reactions that would otherwise be impossible in a biological environment.
Mastering this atomic-level mechanism gives students a profound appreciation for enzymatic precision. If this dangerous radical chemistry were to escape the protective confines of the active site, it would cause massive, indiscriminate cellular damage. The structural architecture of this enzyme evolved to isolate and direct this explosive energy solely toward the targeted synthesis of DNA building blocks.
Slide 12: Regulation of Ribonucleotide Reductase in Nucleotide Biosynthesis
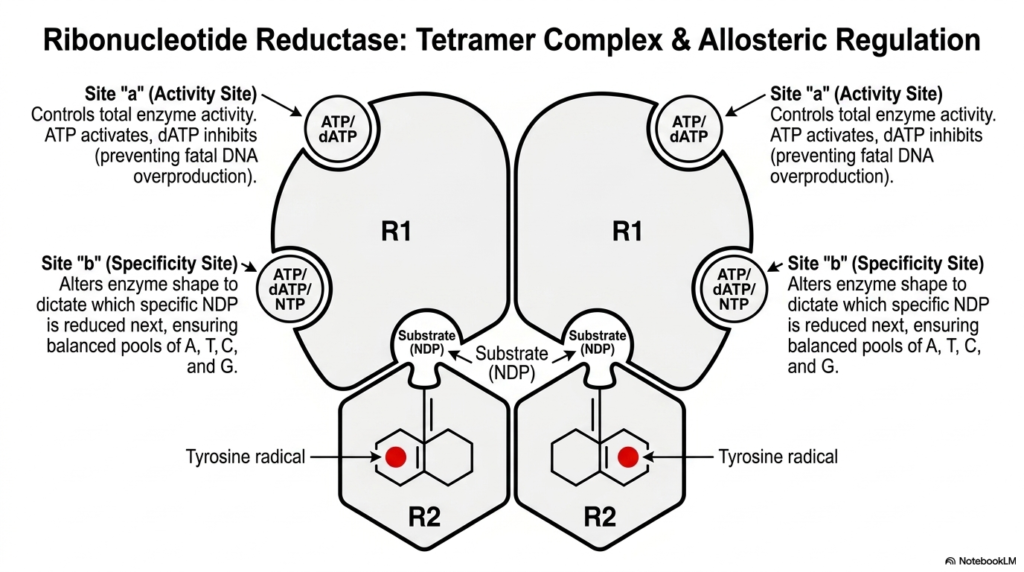
Because ribonucleotide reductase is the sole provider of DNA building blocks, its activity must be perfectly controlled. This slide introduces the multi-layered regulatory system that governs this crucial stage of nucleotide biosynthesis. The enzyme exists as a massive tetramer complex, composed of two R1 subunits and two R2 subunits. The R2 subunits safely house the dangerous tyrosine radicals, while the R1 subunits contain the active sites and critical regulatory control panels.
The enzyme features two distinct allosteric binding sites that dictate its function. Site “a” acts as the primary volume dial, controlling total enzyme activity. When the cell has high levels of ATP, this energetic signal binds to site “a” and turns the enzyme on, indicating the cell is ready to divide. Conversely, high levels of dATP act as a universal off-switch, halting nucleotide biosynthesis to prevent a fatal, toxic overproduction of DNA precursors.
Beyond simply turning on and off, the enzyme must also ensure the genetic alphabet is produced in equal ratios. Site “b” functions as the specificity site. When different nucleotides bind here, they physically alter the shape of the entire enzyme complex. This shape-shifting dictates exactly which specific base will be processed next in the active site. This guarantees a perfectly balanced pool of A, T, C, and G.
This dual-control system prevents catastrophic mutations during DNA replication. If a cell were to produce too much of one specific base, the DNA polymerase would inevitably make lethal copying errors. For biochemistry students studying nucleotide biosynthesis, this enzyme stands as the ultimate paradigm of complex allosteric regulation and structural adaptability.
Understanding these specific allosteric binding sites provides deep insight into complex metabolic feedback loops. The enzyme is constantly tasting the cellular environment, adjusting its internal machinery based on the exact concentrations of available molecules. This level of sophisticated biological engineering ensures that the colossal energy investment required for cellular division is never wasted on unbalanced genetic replication.
Slide 13: Clinical Pathologies of Nucleotide Biosynthesis
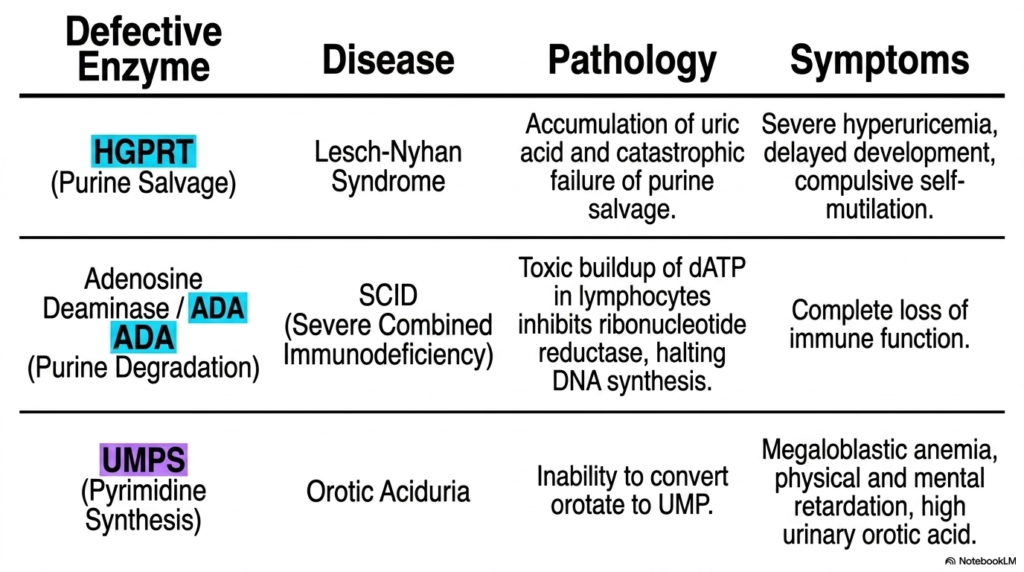
The study of metabolic pathways ultimately culminates in understanding human disease. This impactful slide connects the abstract biochemical pathways of nucleotide biosynthesis directly to severe clinical pathologies. When specific enzymes within these complex networks fail due to genetic mutations, the biological consequences are catastrophic. The table highlights three incredibly important enzyme deficiencies that invariably appear on medical board examinations.
The first pathology results from a failure in the purine salvage pathway. A defective HGPRT enzyme completely halts the recycling process, leading to an extreme accumulation of uric acid. This causes Lesch-Nyhan Syndrome, a devastating disease characterized by severe hyperuricemia, delayed development, and compulsive self-mutilation. This tragically illustrates why the salvage mechanisms in nucleotide biosynthesis are biologically indispensable, particularly for the developing central nervous system.
The second highlighted condition involves a breakdown in purine degradation. When the enzyme Adenosine Deaminase is defective, toxic levels of dATP build up inside white blood cells. This massive surplus allosterically shuts down ribonucleotide reductase, entirely halting new DNA synthesis. The clinical result is Severe Combined Immunodeficiency (SCID), where the patient suffers a complete and fatal loss of immune function.
Finally, the slide addresses a failure in pyrimidine nucleotide biosynthesis. A mutation in the bifunctional UMPS enzyme prevents the essential conversion of orotate to UMP. This molecular roadblock results in Orotic Aciduria. Patients present with severe megaloblastic anemia, physical and mental retardation, and dangerously high levels of orotic acid in their urine. This single slide effectively bridges the gap between basic biochemistry and critical clinical diagnostics.
By examining these specific metabolic failures, students transition from simple memorization to complex clinical reasoning. Recognizing how a single missing protein can trigger widespread, systemic devastation underscores the fragile precision of human metabolism. These tragic pathologies serve as powerful reminders that the intricate enzymatic cascades studied in the classroom are the literal lifelines of human health and development.
Slide 14: Pharmacotherapy Map of Nucleotide Biosynthesis
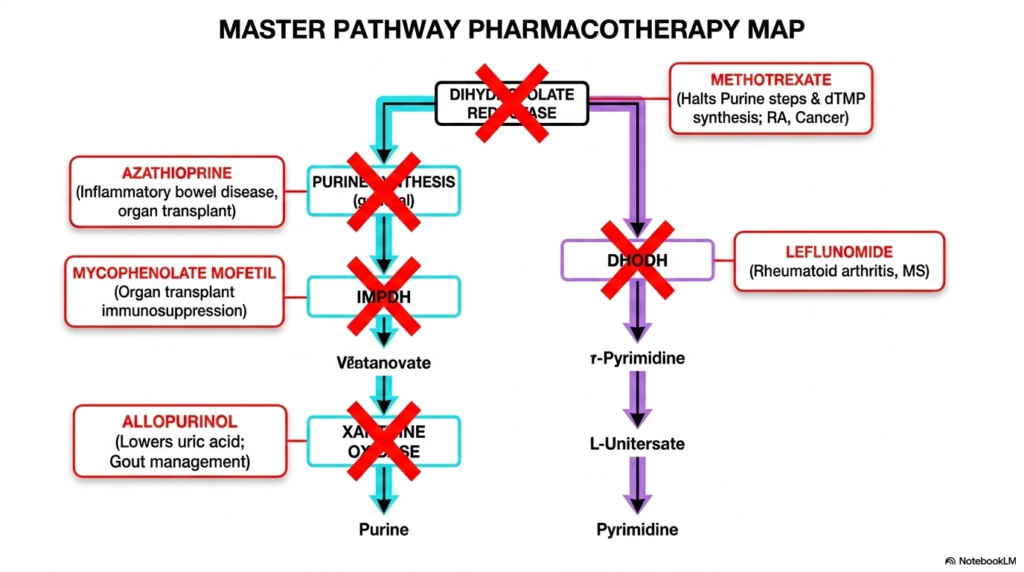
The grand finale of this presentation bridges the gap between complex biochemistry and life-saving medical treatments. This master pharmacotherapy map visualizes how the pathways of nucleotide biosynthesis are deliberately targeted by powerful medications. Because cancer cells and hyperactive immune cells rely heavily on rapid DNA production, intentionally sabotaging these exact metabolic routes is a cornerstone of modern medicine.
The diagram highlights several crucial drug interventions. Methotrexate acts as a profound metabolic poison by inhibiting dihydrofolate reductase. This eliminates the active folate required for purine construction, effectively halting nucleotide biosynthesis and stopping tumor growth. Similarly, Leflunomide targets the pyrimidine pathway by actively destroying the function of the mitochondrial enzyme DHODH, providing immense relief for patients suffering from rheumatoid arthritis.
Other medications target specific branches to manage organ transplants and autoimmune diseases. Mycophenolate Mofetil strategically inhibits IMP dehydrogenase, starving immune cells of the specific guanine precursors they desperately need to multiply. Meanwhile, Azathioprine serves as a more general inhibitor of early purine synthesis. These drugs prove that an intimate understanding of nucleotide biosynthesis is absolutely required to master modern clinical pharmacology.
Finally, the map illustrates the treatment of metabolic waste buildup. By deploying Allopurinol, physicians can intentionally block the enzyme xanthine oxidase. This pharmacological intervention halts the production of dangerous uric acid, serving as the primary management strategy for severe gout. This concluding slide proves to medical students that the complex molecular pathways they study are the exact targets they will manipulate to save patient lives.
This strategic overview synthesizes the entire presentation. It transforms an abstract network of enzymes and chemical intermediates into a functional tactical map used daily by oncologists and rheumatologists. By recognizing the specific vulnerabilities within these metabolic pathways, future healthcare professionals are empowered to understand not just what a drug does, but exactly how and why it works at the molecular level.
Please read our Content Disclaimer Statement.
Check out our social media channels: