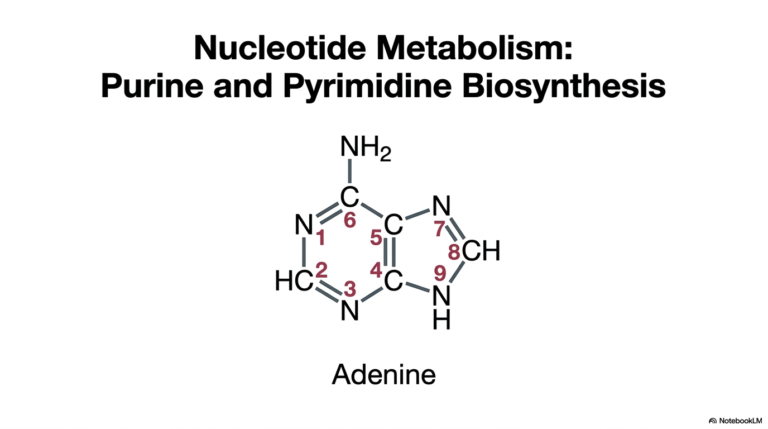
2. Biomolecules Functional Groups: The Cellular Blueprint
Have you ever stopped to marvel at how a microscopic, seemingly chaotic soup of chemicals inside your cells organizes itself to power a beating heart, encode a lifetime of memories, or fight off a viral infection? Our bodies are essentially the universe’s most advanced, highly efficient biological Lego sets, painstakingly constructed from remarkably simple chemical building blocks. The core purpose of this chapter is to deconstruct these fascinating biological blueprints. We are going straight to the foundations of biochemistry to understand exactly how basic atomic elements scale up into the massive, high-energy molecular engines that keep us alive, with a special focus on the ultimate metabolic powerhouse: Acetyl-Coenzyme A.
Here is your comprehensive breakdown of the fundamental rules of biomolecular architecture.
1. Biomolecular Architecture
Welcome to the incredible molecular machinery that sustains human life. This opening slide introduces us to the foundational study of biomolecular architecture, focusing specifically on functional groups and complex coenzymes. These are the tiny, highly specific biochemical tools that cellular proteins and enzymes rely on to do their jobs. Rather than viewing cellular biology as a messy, unpredictable collection of billions of individual atoms, we need to look at it as a beautifully organized, modular system. Just like a master architect uses a set of standardized blueprints and fundamental raw materials to construct a towering skyscraper, the living cell uses strict biochemical principles to build everything from your DNA strands to the metabolic engines that burn the food you eat. By analyzing these critical biochemical concepts, medical and college students can truly understand how simple elements form the complex, life-sustaining molecules that define human physiology. Mastering this architectural view is essential because modern medicine, pharmacology, and targeted drug design rely entirely on an understanding of these specific atomic shapes and molecular structures.
2. The Modular Principle of Life
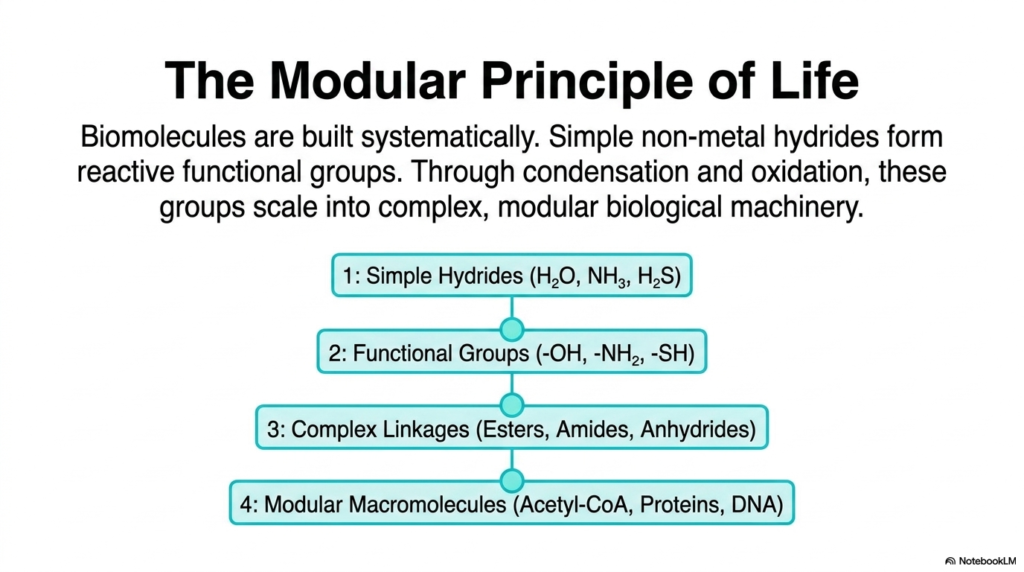
When you look at massive, complex biomolecules like human DNA, towering protein structures, or dense lipid bilayers, it is incredibly easy to feel overwhelmed by their sheer size. However, life is phenomenally efficient and relies heavily on what biochemists call the modular principle. Biomolecules are not built haphazardly; they are constructed systematically from the ground up. This slide illustrates how we transition from incredibly simple, nonmetallic hydrides—such as water, ammonia, or hydrogen sulfide—to highly reactive functional groups. Through highly predictable chemical reactions, specifically condensation (bringing pieces together by removing water) and oxidation (swapping electrons and hydrogens), these small functional groups scale up. They form complex chemical linkages, eventually creating the massive, modular biological machinery necessary for cellular life, such as Acetyl-CoA, proteins, and DNA. Think of it like assembling a luxury vehicle on a factory line: you don’t start with a fully formed car. You start with basic raw materials, forge them into specific functional parts, link those parts together, and eventually assemble the complex macromolecule.
3. Derivatives of Non-Metal Hydrides
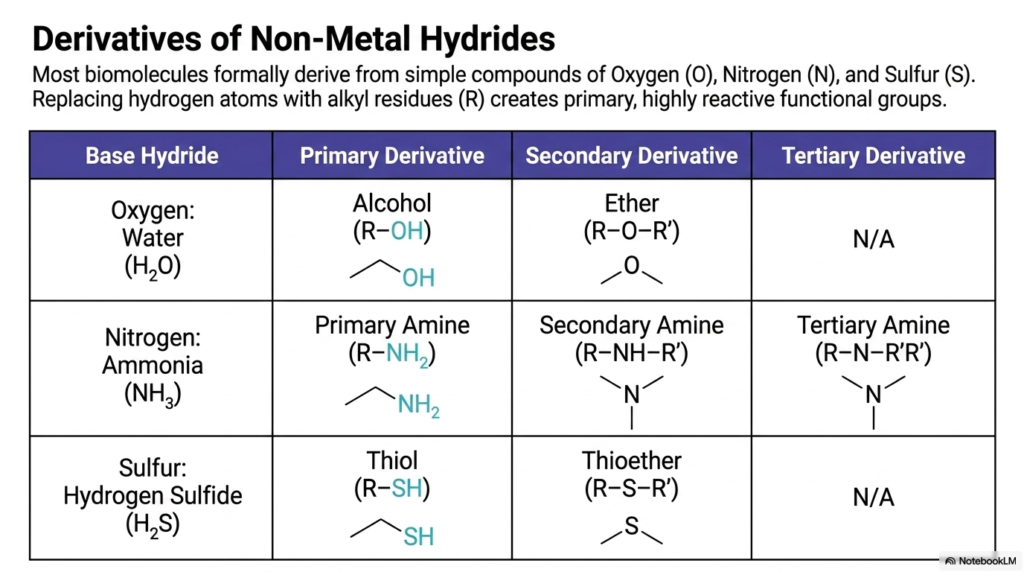
To truly master the language of biochemistry, you have to understand the basic elemental ingredients of the human cell. Most biomolecules formally derive from incredibly simple compounds made of Oxygen (O), Nitrogen (N), and Sulfur (S). This slide serves as your molecular periodic table of functional groups. By simply taking a base non-metal hydride—such as water (H2O), ammonia (NH3), or hydrogen sulfide (H2S)—and replacing its hydrogen atoms with carbon-based alkyl groups (R-groups), the cell creates its primary toolkit. When you replace a hydrogen on water with an alkyl group, you generate an alcohol. Do it twice, and you get an ether. When you systematically modify ammonia, you create primary, secondary, and tertiary amines. Modifying hydrogen sulfide yields highly reactive thiols and thioethers. These modifications aren’t just random chemical tweaks; these functional groups strictly dictate exactly how a molecule will behave, whether it will dissolve easily in your bloodstream, how it will carry an electrical charge, or how tightly it will bind to a cellular receptor.
4. The Oxidation Ladder
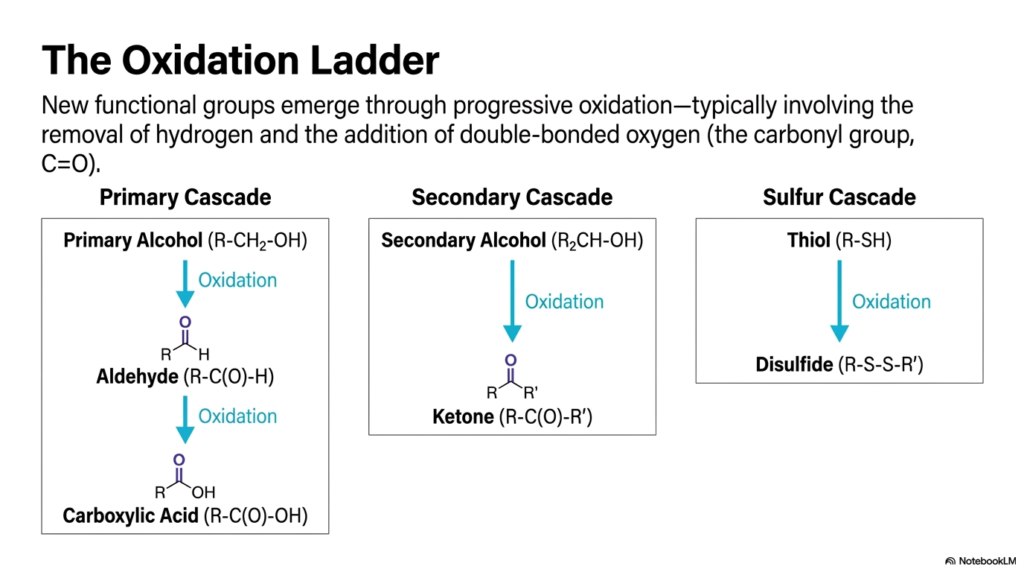
If functional groups are the cell’s basic toolkit, the oxidation ladder is the exact mechanism the cell uses to upgrade its tools. New, highly reactive functional groups emerge through progressive oxidation. This chemical process typically involves the removal of hydrogen atoms and the simultaneous addition of a double-bonded oxygen, famously known in chemistry as a carbonyl group. This slide brilliantly maps out these progressive cascades. In the primary cascade, a relatively stable primary alcohol is oxidized into a highly reactive aldehyde, which can then be further oxidized into a very stable carboxylic acid. Secondary alcohols climb their own unique ladder to become ketones. Down in the sulfur cascade, a thiol is oxidized to form a rigid, stabilizing disulfide bond, which is critical for holding protein structures together. This progressive increase in oxidation states is how cells meticulously control chemical reactivity. It is the exact process your liver uses to break down toxins and how your mitochondria extract energy from carbohydrates.
5. Carbonyl Addition Reactions
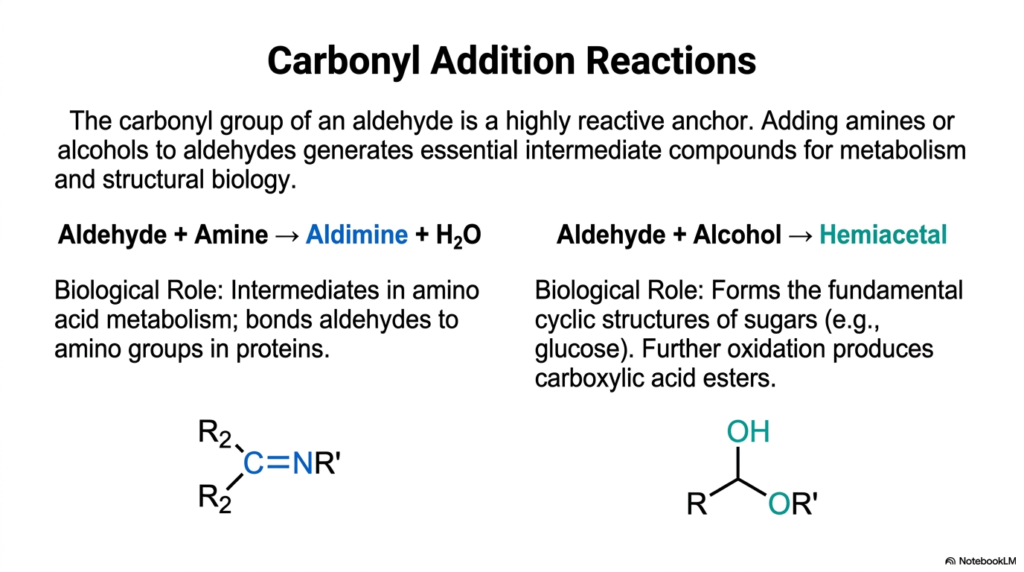
Now we are getting into the actual construction phase of cellular machinery. The carbonyl group—that double-bonded oxygen we just explored on the oxidation ladder—is essentially a highly reactive anchor point for the cell. This slide demonstrates exactly what happens when you combine an aldehyde with other functional groups to build larger structures. When you add an amine to an aldehyde, you generate an aldimine and release a molecule of water. This specific reaction is a crucial intermediate step in amino acid metabolism and helps bind aldehydes to amino groups in proteins. When you add an alcohol to an aldehyde, you form a hemiacetal. These addition reactions are the fundamental ways that human cells snap different biological modules together to create larger, more complex structures. For instance, the formation of a hemiacetal is the exact reaction that allows simple, linear sugars, like glucose, to fold into their fundamental cyclic ring structures, making them ready for cellular energy production.
6. Carboxylic Acid Derivatives
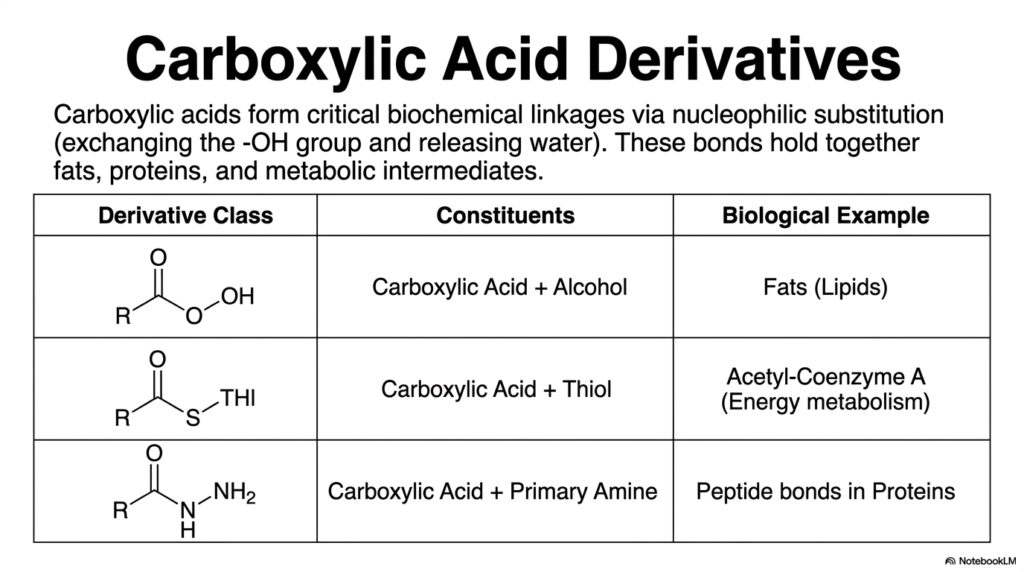
This slide introduces the heavy-duty structural linkages of the biological world. Carboxylic acids form critical biochemical bonds through a process called nucleophilic substitution—essentially swapping out an -OH group and kicking out a molecule of water to bind with a new group. When a carboxylic acid reacts with an alcohol, it forms an ester. This is the exact bond that holds our fat stores (lipids) together. When it reacts with a highly reactive thiol, it forms a high-energy thioester, which is the beating heart of energy metabolism molecules like Acetyl-Coenzyme A. Finally, when a carboxylic acid reacts with a primary amine, it forms an amide. Amide bonds are the famous peptide bonds that string amino acids together to build every single protein in your body. These specific derivatives are the structural steel beams and the high-voltage power lines of the human cell, providing both the stability to hold tissues together and the dynamic energy required to fuel human movement.
7. Phosphorus: The Energy Anchor
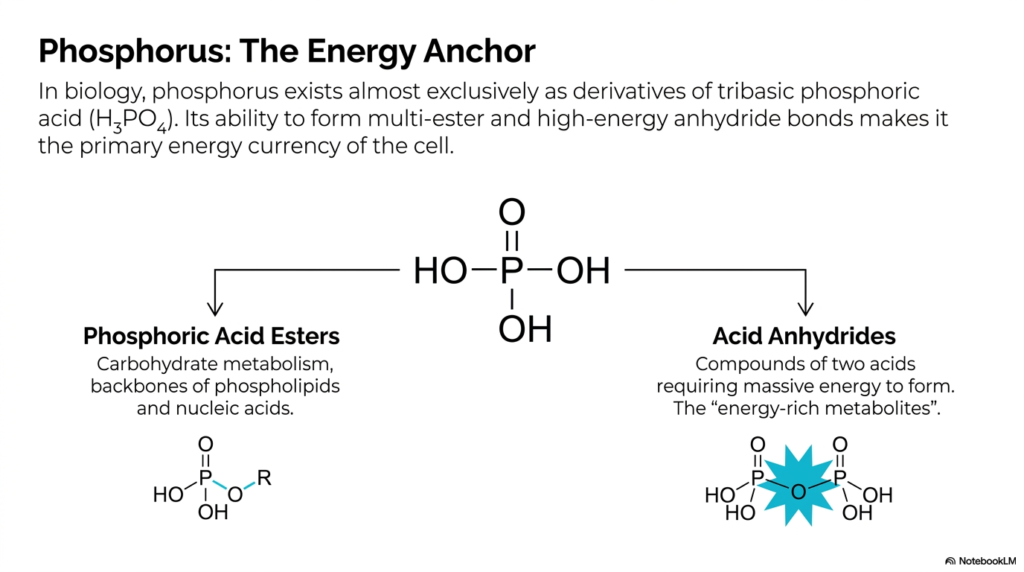
If carbon provides the structural backbone of biological life, phosphorus acts as its high-voltage electricity. In biology, phosphorus exists almost exclusively as derivatives of tribasic phosphoric acid (H3PO4). This slide highlights its two vastly different, yet equally critical, cellular roles. First, it forms phosphoric acid esters. These multi-ester bonds are the rigid, incredibly stable backbones of our DNA strands and the complex phospholipids that make up every single cell membrane in your body. Second, and much more dynamically, phosphorus can form acid anhydrides. Anhydrides are compounds formed by forcing two acids together, a process that requires a massive, intense input of energy. When the cell successfully creates a phosphoric acid anhydride bond, it is essentially compressing a highly volatile molecular spring. These acid anhydride bonds are the universal energy currency of all known life, making them the “energy-rich metabolites” that power everything from brain function to muscle contraction.
8. Modularity via Water: Condensation & Hydrolysis
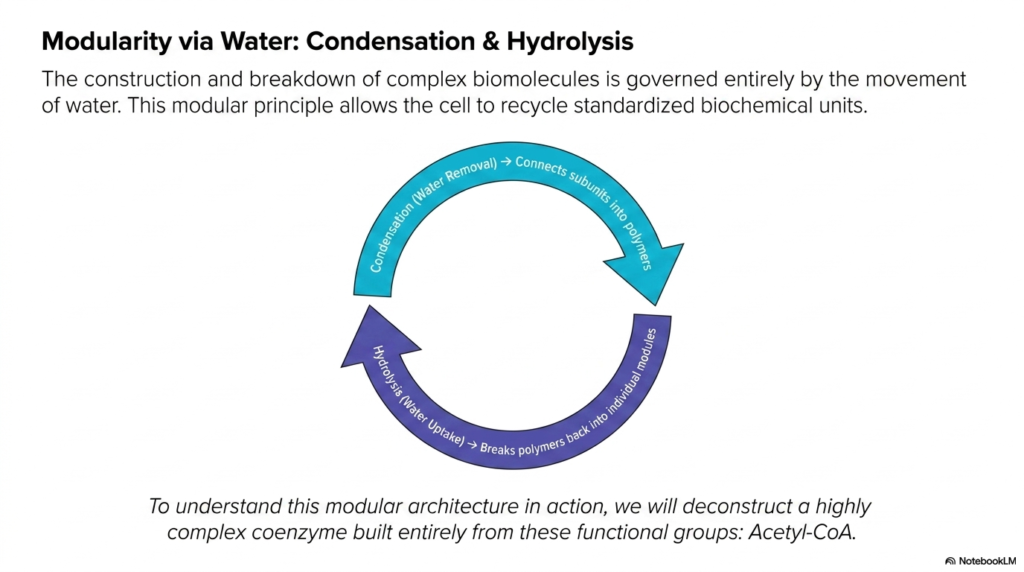
It is a beautiful, elegant paradox that water is both the absolute simplest molecule in our bodies and the master regulator of our most complex biochemical processes. The construction and breakdown of all complex biomolecules are governed entirely by the movement of water. As the cycle on the slide shows, condensation involves the removal of water to join molecular subunits into larger, functional polymers. This is how your body builds muscle tissue or stores complex carbohydrates. Conversely, hydrolysis is the uptake of water to break those massive polymers back down into their individual, standardized chemical modules. This elegant, highly reversible cycle allows the human cell to constantly recycle its biochemical units without wasting precious energetic resources. To truly understand this modular architecture in action, the remainder of our exploration will deconstruct a highly complex coenzyme built entirely by these precise, water-driven functional groups: Acetyl-CoA.
9. Acetyl-CoA: The High-Potential Engine
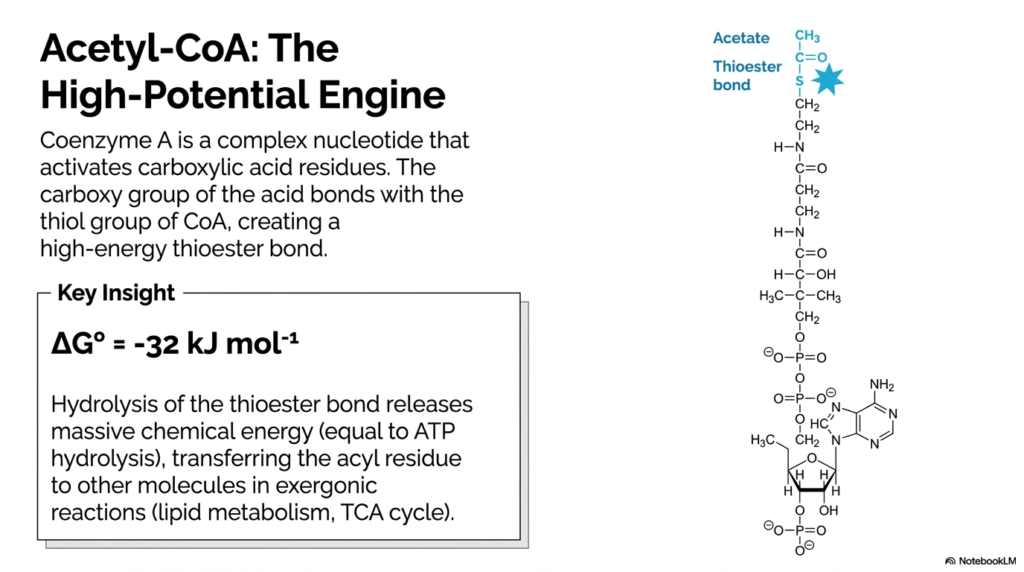
Meet the most important biological molecule you will ever study: Acetyl-Coenzyme A (Acetyl-CoA). This complex nucleotide is the ultimate, high-potential metabolic engine of the cell. Its primary physiological job is to activate carboxylic acid residues so the body can use them. The vital chemical connection occurs when the carboxylate group of an acetate molecule binds directly to the reactive thiol group of Coenzyme A. This creates a high-energy thioester bond. Why is this specific cyan-burst bond so incredibly special? Because the hydrolysis of this thioester bond releases a massive amount of chemical energy—a staggering ΔG° of -32 kJ mol-1. To put that in perspective, this is exactly equal to the explosive energy released by breaking an ATP molecule! This immense power is used to safely transfer the acyl residue to other molecules, driving the vital exergonic reactions that keep our bodies running, such as lipid metabolism and the TCA (Krebs) cycle.
10. Acetyl-CoA Modules 1 & 2: Cysteamine and β-Alanine
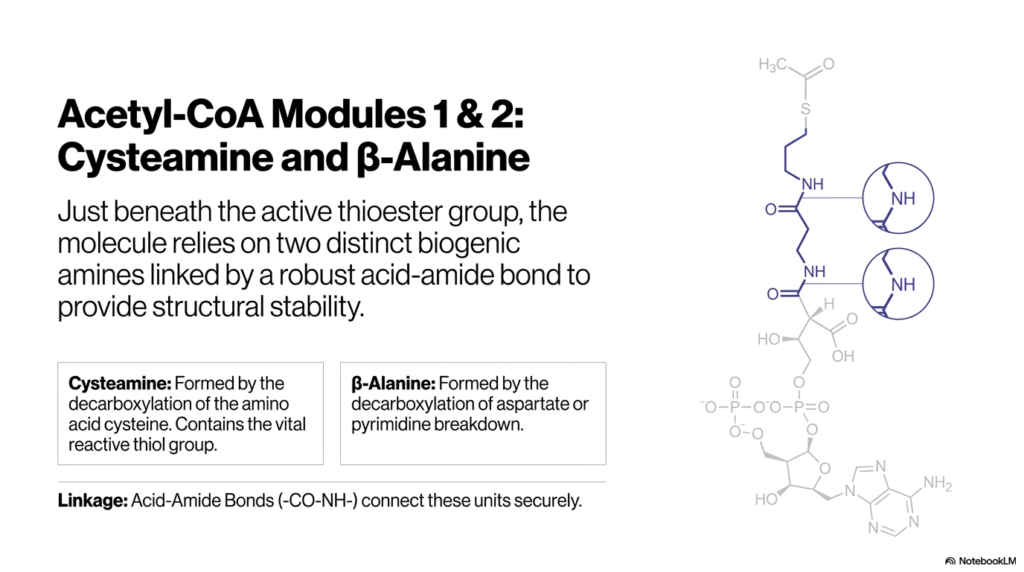
To fully appreciate how Acetyl-CoA functions safely without spontaneously detonating its high-energy payload in the cytoplasm, we have to look beneath the hood at its individual structural modules. Just beneath the active thioester group, the molecule relies on two distinct biogenic amines: Cysteamine and β-Alanine. Cysteamine is formed by the decarboxylation of the amino acid cysteine. Crucially, this module provides the vital reactive thiol group (-SH) that forms the high-energy thioester bond at the top of the molecule. Directly below it is β-Alanine, which is formed from the metabolic breakdown of aspartate or pyrimidines. These two essential modules are linked together by a robust, highly stable acid-amide bond (-CO-NH-). This strong, peptide-like linkage ensures that while the very tip of the molecule is violently reactive and ready to transfer energy, the structural “arm” holding it remains completely intact and secure during the chaos of cellular metabolism.
11. Acetyl-CoA Module 3: Pantoinate & Chirality

Moving further down the complex Acetyl-CoA structure, we encounter a fascinating module that introduces a strict environmental and dietary dependency into human biology: Pantothenate. Linked to the β-Alanine above it via another highly stable acid-amide bond, pantoinate brings two critical features to the table. First, it contains a stereospecific chiral center (marked by a purple asterisk). This means the molecule has a very specific 3D “handedness,” and in natural Coenzyme A, only the (R)-pantoinate enantiomer is biologically functional. Second, human metabolism completely lacks the enzymes to synthesize pantoinate from scratch. It must be acquired from our diet, combined with β-Alanine, as Pantothenate—widely known as Vitamin B5. Furthermore, this entire upper segment of the molecule is so structurally useful that it often functions independently in specific proteins, serving as a standalone 4′-phosphopantetheine arm in enzymes such as fatty acid synthase.
12. Acetyl-CoA Modules 4, 5, 6: The Phosphoric Bridge
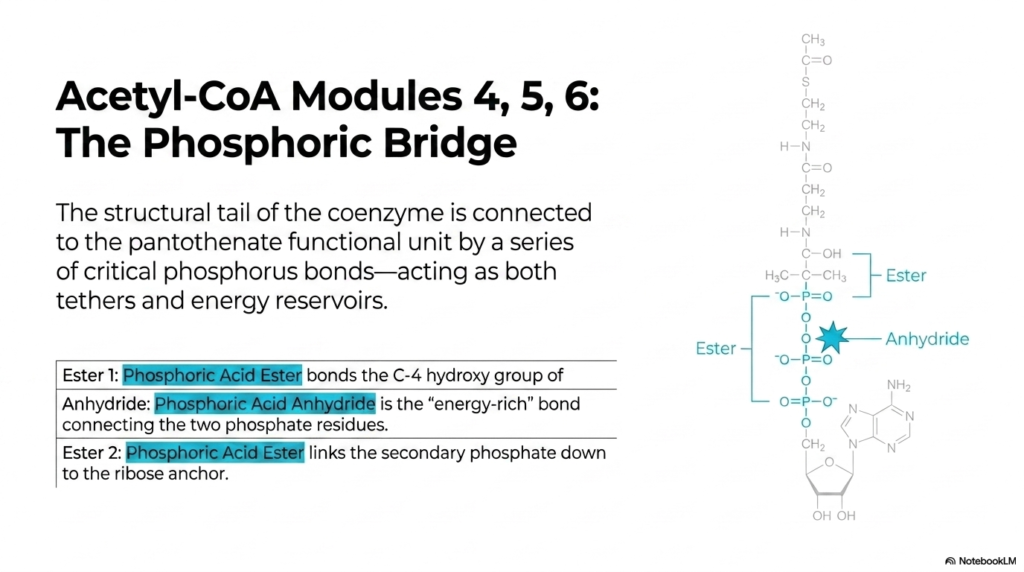
Connecting the highly active, vitamin-derived “business end” of Acetyl-CoA to its deep structural base requires a highly engineered molecular tether. This slide deconstructs the structural Phosphoric Bridge. The bridge consists of a precise, alternating sequence of vital phosphorus bonds that act as both tethers and energy reservoirs. First, a Phosphoric Acid Ester (Ester 1) tightly bonds the C-4 hydroxyl group of the pantothenate tail. Next, right in the middle, sits a high-energy Phosphoric Acid Anhydride bond connecting two phosphate residues. Finally, a second Phosphoric Acid Ester (Ester 2) links the secondary phosphate securely to the ribose anchor below. This bridge doesn’t just passively hold the giant molecule together; it acts as a flexible, energy-rich shock absorber. It ensures the molecule has the exact geometric length and flexibility required to reach deep into the active sites of complex metabolic enzymes.
13. Acetyl-CoA Modules 7 & 8: The Adenosine Anchor
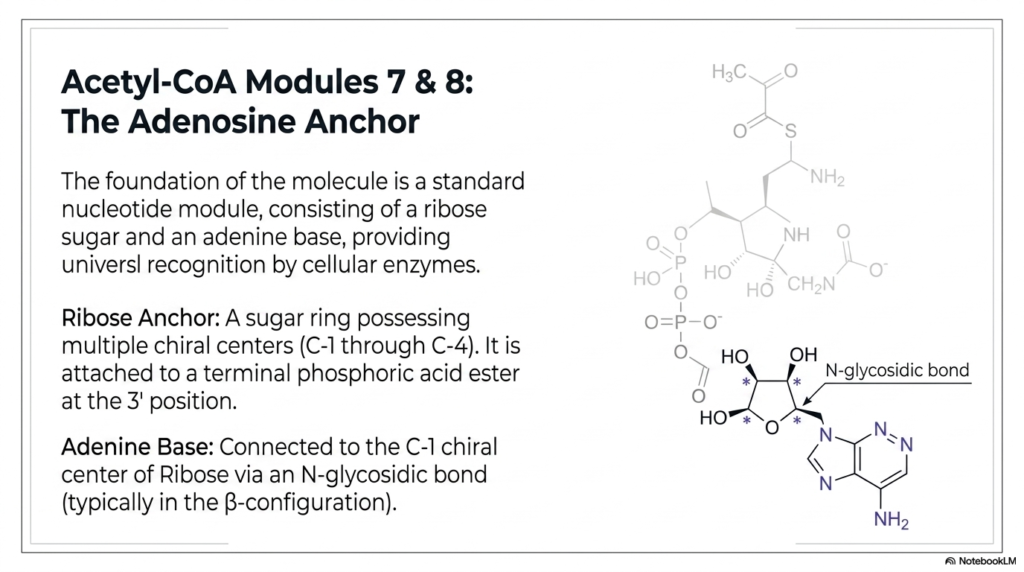
Every massive, complex biological machine needs a rock-solid foundation, and for Acetyl-CoA, that foundation is the Adenosine Anchor. The absolute bottom of the molecule is a standard nucleotide module, structurally identical to the fundamental building blocks of human RNA. It consists of two main parts. First is the Ribose Anchor, a sugar ring possessing multiple chiral centers (C-1 through C-4) that dictate its exact 3D orientation. It is attached to a terminal phosphoric acid ester at the 3′ position. Second is the Adenine Base, which is connected to the C-1 chiral center of the Ribose via a sturdy N-glycosidic bond (typically in the β-configuration). Why use a standard nucleotide to anchor a metabolic energy-transfer molecule? Because adenine provides universal recognition. It acts as a cellular barcode, allowing a wide variety of cellular enzymes to instantly recognize, bind, and utilize the coenzyme efficiently.
14. The Complete Architectural Blueprint: Acetyl-CoA
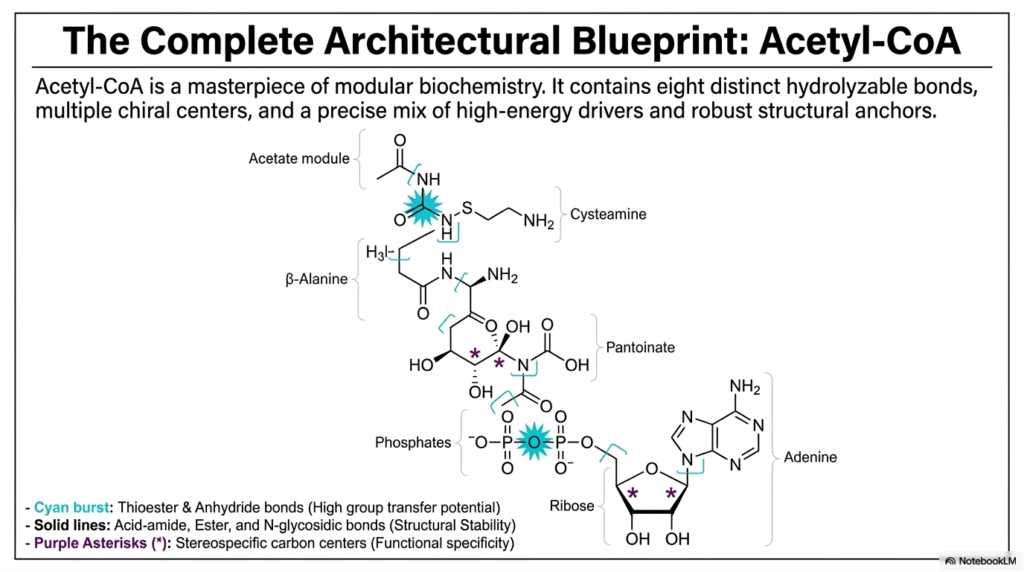
When we finally assemble all of the individual modules we have discussed, the complete architectural blueprint of Acetyl-CoA is revealed as an absolute masterpiece of modular biochemistry. It is abundantly clear that this is not just a random jumble of carbon and nitrogen atoms; it is a highly engineered biological machine. The completed coenzyme contains eight distinct hydrolyzable bonds, multiple chiral centers (highlighted by purple asterisks) that require highly specific 3D geometry, and a precisely calculated mix of high-energy metabolic drivers alongside remarkably robust structural anchors. The cyan bursts represent the explosive thioester and anhydride bonds that provide high group-transfer potential. The solid lines represent the unbreakable acid-amide, ester, and N-glycosidic bonds that provide structural stability. This delicate, perfect balance of explosive energy transfer and rigid structural stability is exactly what allows Acetyl-CoA to safely ferry massive amounts of chemical energy throughout the human cell.
15. From Elements to Engines
This final concept flawlessly synthesizes the overarching thesis of biomolecular architecture: truly understanding biomolecules requires looking far past raw, intimidating chemical formulas to see the underlying, elegant architectural logic of the cell. We have seen that chemical reactivity is never random but is strictly defined by predictable derivative functional groups. We have observed how immense molecular complexity arises from highly predictable, simple chemical reactions such as cellular oxidation and condensation. Above all, we have proven that biological function is inherently modular. The human cell takes standardized, basic chemical units—such as simple amines, essential dietary vitamins, and standard nucleotides—and seamlessly mixes, matches, and bonds them to create highly specific biological machines capable of sustaining human life. Understanding this modular paradigm is the key to mastering biochemistry, unlocking the secrets of cellular metabolism, and advancing modern medicine.
Check out our social media channels:
Please read our Content Disclaimer Statement.