4. Bioenergetics: How Cells Generate and Harness Energy
Picture this: you are sprinting to catch a bus, your heart is pounding, and your muscles are burning. In that exact moment, billions of your cells are frantically burning through molecular fuel to keep you moving. But how exactly does a microscopic blob of water and proteins generate the sheer mechanical force needed to propel your body forward? That is the beautiful, complex puzzle we are going to solve today.
This chapter is your ultimate guide to the physics of life. We are going straight into the heart of cellular energy to understand the thermodynamic and kinetic engines that make biological work possible. Whether you are a college student cramming for a biochemistry final or a medical student trying to make sense of metabolic pathways, understanding these fundamental driving forces is essential. We will break down everything from Gibbs free energy to reaction speeds, translating complex equations into the living, breathing mechanisms that power your physiology.
Slide 1: Introduction to Bioenergetics
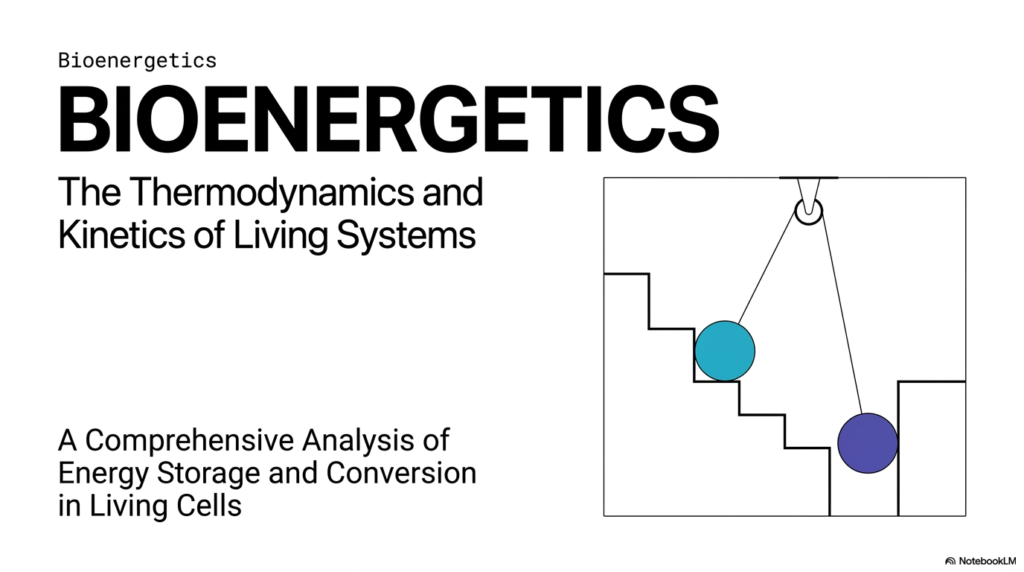
Have you ever stopped to wonder how the human body takes the food you eat and magically turns it into the energy needed to run a marathon, study for a grueling pharmacology exam, or simply keep your heart beating while you sleep? That is the fascinating realm of Bioenergetics. Welcome to the foundational physics of life itself. In this opening slide, we are introduced to the overarching theme of Bioenergetics, which elegantly bridges the gap between cold, hard thermodynamics and the dynamic kinetics of living systems.
Think of a human cell as a bustling, microscopic metropolis. It requires a ceaseless, uninterrupted supply of power to keep the lights on, transport vital cargo across membranes, and rebuild structural damage. Bioenergetics is the dedicated study of exactly how that biological power is acquired, stored, and utilized without breaking the fundamental laws of physics. For college and medical students, mastering Bioenergetics is not just about memorizing random metabolic pathways on a whiteboard; it is about deeply understanding the ‘why’ behind all of cellular life. Why does the breakdown of ATP drive muscle contraction? Why do certain chemical reactions happen instantly while others require specific enzymes to proceed?
This comprehensive slide deck serves as an in-depth analysis of energy storage and conversion in living cells. We will soon explore the delicate balance between thermodynamics—the absolute rules that dictate whether a reaction can occur—and kinetics—the rules that dictate how fast it occurs. By understanding the core principles of Bioenergetics, you gain the fundamental toolkit required to diagnose metabolic diseases, understand pharmacology at a molecular level, and appreciate the incredible elegance of human physiology. Keep in mind that the principles of energy storage govern every single process in a living organism.
Slide 2: The Anatomy of Work in Bioenergetics
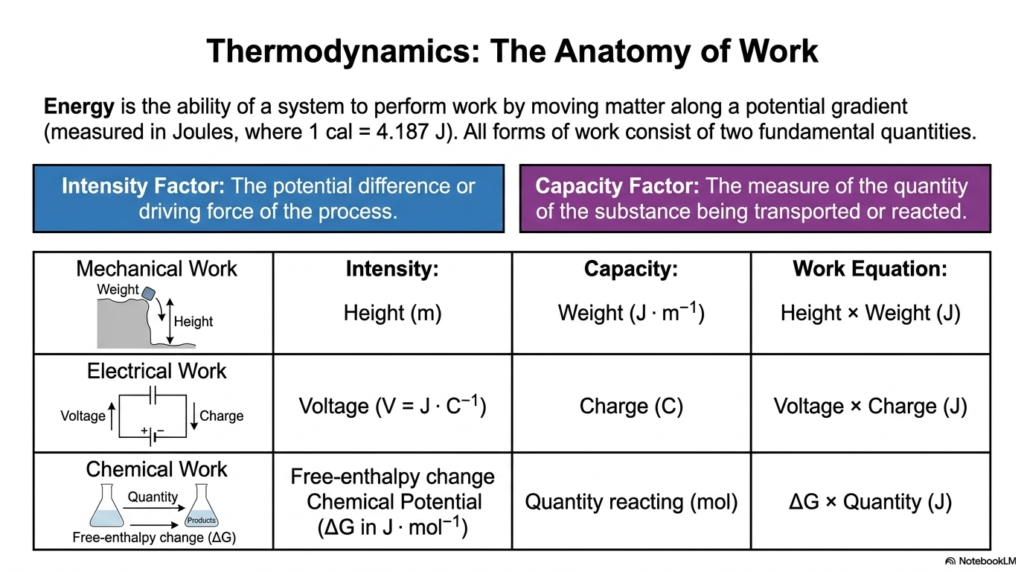
When we talk about biological work, we have to borrow some concepts from physics and apply them to the messy reality of biology. This slide breaks down the core components of energy in Bioenergetics. In physics, energy is simply the ability of a system to perform work by moving matter along a potential gradient. However, all forms of work—whether mechanical, electrical, or chemical—can be split into two fundamental factors: the Intensity Factor and the Capacity Factor.
The Intensity Factor is the driving force or the potential difference. Think of it as the steepness of a hill. The Capacity Factor is the actual amount of stuff being moved. In mechanical work, the intensity is the height of the drop, and the capacity is the weight of the object. In electrical work, intensity is voltage, and capacity is electrical charge. But for those studying Bioenergetics, our main focus is chemical work. Here, the intensity factor is the change in free enthalpy, also known as the chemical potential (measured in Joules per mole). The capacity factor is the number of reacting molecules.
Why does this matter for your cellular biology exams? Because your cells are constantly performing chemical work to stay alive. The principles of Bioenergetics dictate that every time a molecule is synthesized or an ion is pumped across a cell membrane, the cell must expend energy. By understanding that chemical work is simply the product of the chemical driving force (free enthalpy) and the amount of substance reacting, you can start to see metabolism as a series of calculated physical transactions. Every heartbeat and every neuron firing is just a manifestation of these intensity and capacity factors working together in perfect harmony.
Slide 3: Energetic Coupling and Bioenergetics
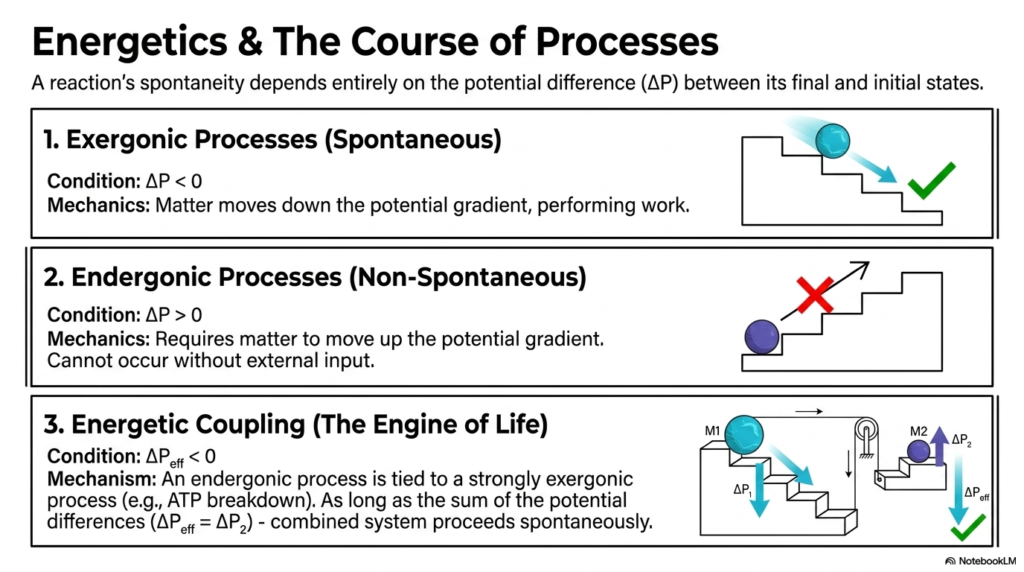
Now that we know what chemical work is, we have to ask: why do some reactions happen on their own while others stubbornly refuse to budge? This brings us to the most vital concept in Bioenergetics: reaction spontaneity and energetic coupling. A reaction’s spontaneity depends entirely on the potential difference between its final and initial states.
If a process is exergonic, it means matter moves down the potential gradient. It loses energy, meaning the change in potential is less than zero. These reactions occur spontaneously and perform work, much like a ball rolling effortlessly down a staircase. On the flip side, endergonic processes require matter to move up the potential gradient. These are absolutely non-spontaneous. You cannot roll a ball up a staircase without physically pushing it. In the context of Bioenergetics, your cells constantly need to perform endergonic tasks, such as building large DNA molecules or pumping calcium into the sarcoplasmic reticulum. So, how do they manage the impossible?
The secret is Energetic Coupling, aptly dubbed the engine of life. The cell brilliantly pairs a heavily endergonic (non-spontaneous) process with a massively exergonic (spontaneous) process. The most famous example in Bioenergetics is the breakdown of ATP. ATP hydrolysis releases so much energy that it acts as a heavy weight pulling a lighter weight up a pulley system. As long as the combined sum of the potential differences is negative, the entire coupled system will proceed spontaneously. This is why ATP is the universal currency of the cell. Without energetic coupling, complex life would literally grind to a halt, as your cells would lack the mechanical force to drive the uphill battles of metabolism.
Slide 4: Chemical Equilibrium in Bioenergetics
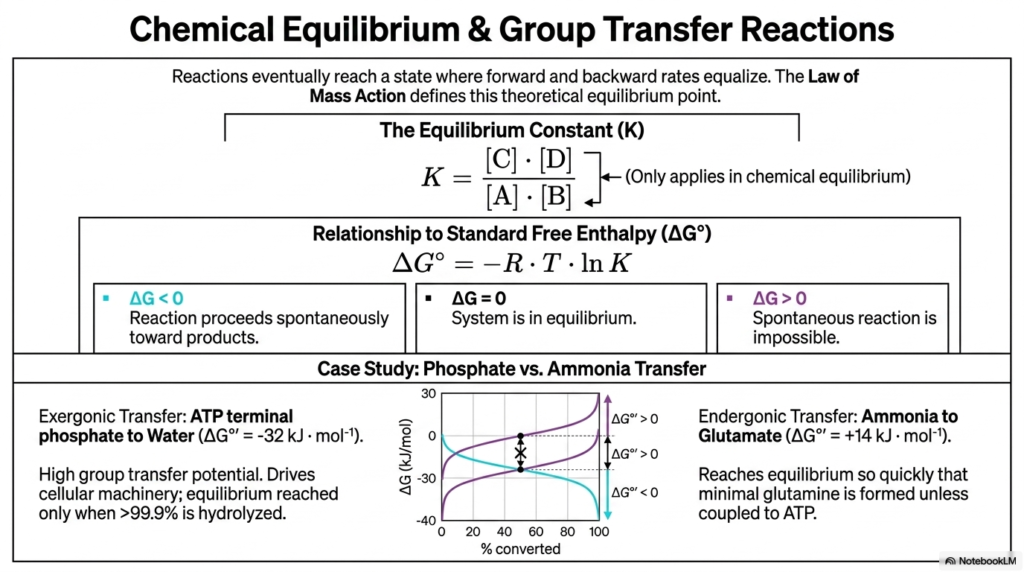
One of the greatest illusions in chemistry is the idea that reactions only go in one direction. In reality, reactions eventually reach a state where the forward and backward rates equalize. This slide introduces chemical equilibrium, a deeply important concept for understanding Bioenergetics. According to the Law of Mass Action, this theoretical point is defined by the Equilibrium Constant (K), which is simply the ratio of products to reactants.
The relationship between this constant and the standard free enthalpy (ΔG) dictates the direction of life. If ΔG is less than zero, the reaction proceeds spontaneously to form products. If it is greater than zero, the reaction is impossible without outside help. If ΔG equals zero, the system is in perfect equilibrium. However, in the world of Bioenergetics, total equilibrium actually means cell death. Living cells must constantly maintain a state of disequilibrium to keep performing work.
Let us look at the case study comparing phosphate transfer to ammonia transfer. The transfer of the terminal phosphate from ATP to water is highly exergonic. ATP has a massive group-transfer potential, strongly favoring the forward reaction until more than 99.9% of it has been hydrolyzed. Conversely, transferring an amino group from ammonia to glutamate to form glutamine is endergonic. On its own, it reaches equilibrium so fast that almost no glutamine is made. To survive, the body uses the principles of Bioenergetics to couple this stubborn ammonia transfer to the explosive power of ATP hydrolysis. Understanding how these equilibrium points shift allows medical students to grasp how drugs, toxins, and metabolic diseases alter the balance of life.
Slide 5: Redox Potential and Bioenergetics
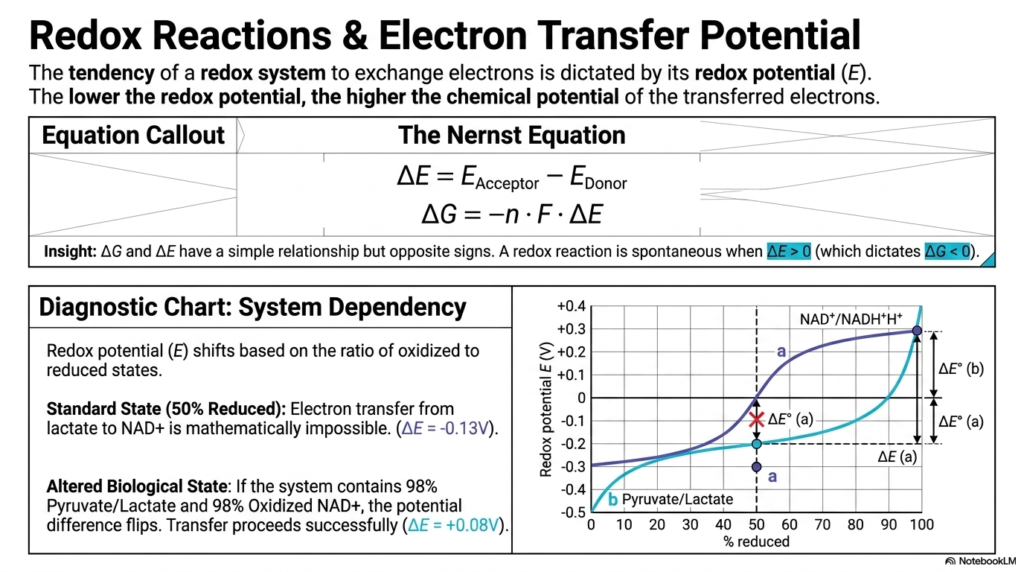
Beyond transferring phosphate groups, cells survive by transferring electrons. If you want to understand the electron transport chain in the mitochondria, you must first master the principles of redox reactions shown on this slide. In Bioenergetics, the tendency of a biological system to exchange electrons is dictated by its redox potential (E). The golden rule here is simple: the lower the redox potential, the higher the chemical potential of the transferred electrons.
To calculate the exact driving force of these electrons, we use the Nernst Equation. It shows us that the change in free energy (ΔG) is directly proportional to the change in redox potential, but with an opposite sign. A positive change in redox potential means a spontaneous, negative change in free energy. This is a crucial concept in Bioenergetics because it explains how your cells generate the voltage that sustains life.
Let’s look at the diagnostic chart provided on the slide. Under standard conditions (where species are 50% reduced), transferring an electron from lactate to NAD+ is mathematically impossible because the potential difference is negative. But living cells are not standard test tubes. In an altered biological state—where there is a massive surplus of pyruvate and oxidized NAD+—the potential difference completely flips! The transfer now proceeds successfully. This perfectly illustrates why Bioenergetics is so vital for future physicians to understand. By constantly shifting the concentrations of oxidized and reduced molecules, your cells manipulate redox potentials to drive otherwise impossible reactions to extract energy.
Slide 6: Acid-Base Reactions in Bioenergetics
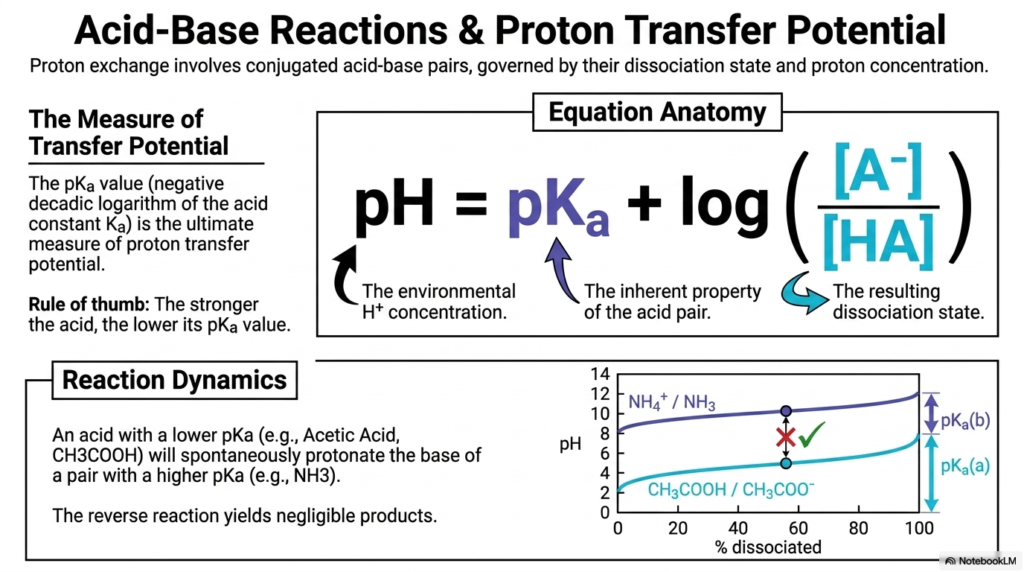
While electrons are shuttled around to generate power, protons (hydrogen ions) also play a massive role in cellular homeostasis. This slide breaks down acid-base reactions and the concept of proton transfer potential, another core pillar of Bioenergetics. Whenever we talk about proton exchange, we are looking at conjugated acid-base pairs. Their behavior is strictly governed by their dissociation state and the surrounding environmental proton concentration, commonly known as pH.
The ultimate measure of a molecule’s proton transfer potential is its pKa value. The pKa is the inherent property of the acid pair, representing the negative decadic logarithm of the acid constant. As a fundamental rule of thumb in Bioenergetics, the stronger the acid, the lower its pKa value. We calculate the resulting pH of a biological environment using the Henderson-Hasselbalch equation anatomy shown on the slide, mapping how the inherent pKa interacts with the ratio of dissociated to intact acid.
To see this in action, observe the reaction dynamics chart. An acid with a drastically lower pKa, such as acetic acid, will naturally and spontaneously donate a proton to the base of a pair with a higher pKa, such as ammonia. The reverse reaction is practically non-existent, yielding negligible products. Why should a medical or biochemistry student care about this aspect of Bioenergetics? Because every enzyme in your body relies on precise protonations to function. Medications are absorbed based on their pKa relative to the stomach and intestinal pH. The cell relies entirely on proton gradients to drive ATP synthesis. Mastering the dynamics of proton transfer is absolutely essential to understanding overall metabolic balance.
Slide 7: Enthalpy and Heat in Bioenergetics

When chemical bonds form or break, heat is invariably exchanged. This brings us to Enthalpy (ΔH), a critical physical concept explored in this slide. In the realm of Bioenergetics, we measure this heat exchange at a constant pressure. Chemical reactions are broadly categorized into two distinct types based on heat flow: exothermic and endothermic.
Exothermic reactions are processes in which the system loses heat to its surroundings, resulting in a negative ΔH. In a biological context, this heat release closely matches the overall free-energy change during the breakdown and oxidation of dietary nutrients. Endothermic reactions, conversely, pull heat in from the environment, resulting in a positive ΔH. A major goal of Bioenergetics is to calculate exactly how much chemical work can be extracted from this heat flow.
To measure this practically, scientists use a device called a pressurized physical calorimeter. If you want to know the maximum chemical work supplied by a piece of food, you can literally burn a weighed amount of it in a pressurized vessel. As the food is entirely oxidized by oxygen into carbon dioxide and water, the released enthalpy heats the surrounding water bath. Because we know that 1 kilojoule of heat warms 1 liter of water by 0.24 degrees Celsius, we can calculate the exact energy potential of the food.
Understanding this facet of Bioenergetics allows students to connect the abstract concept of “calories” on a nutrition label to the physical laws of thermodynamics. It proves that the human body is essentially a highly regulated combustion engine powered by biochemical heat exchange.
Slide 8: Entropy and Order in Bioenergetics
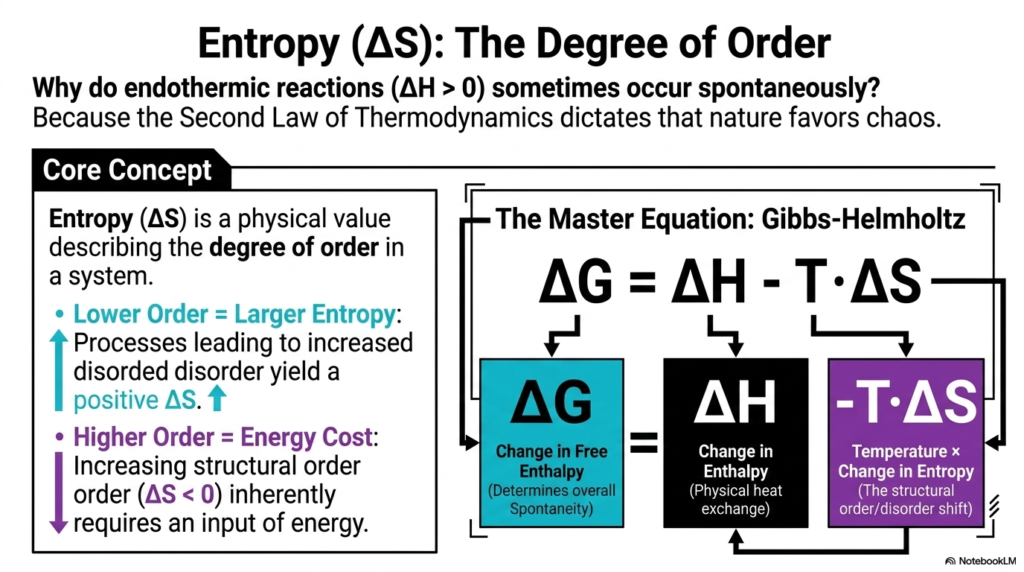
If exothermic reactions release heat and favor spontaneity, why do some endothermic reactions—which absorb heat—still happen spontaneously? This fascinating paradox introduces us to Entropy (ΔS), the ultimate measure of chaos and disorder. As taught in Bioenergetics, the Second Law of Thermodynamics strictly dictates that nature universally favors chaos.
Entropy is a physical value describing the degree of order within any given system. If a biochemical process leads to a lower degree of structural order—meaning increased disorder—it results in a positive change in entropy. Conversely, forcing molecules into a higher state of order inherently requires a massive input of energy, resulting in a negative entropy shift. In Bioenergetics, we synthesize these competing forces using the master equation of life: the Gibbs-Helmholtz equation.
This crucial equation states that the overall change in Free Enthalpy (ΔG), which determines true spontaneity, equals the change in Enthalpy (heat) minus the product of Temperature and the change in Entropy (order). This means that a reaction can still be spontaneous even if it absorbs heat, provided it generates enough molecular chaos to offset the energy cost. For college students studying Bioenergetics, this equation is the skeleton key to understanding biochemistry. It explains why proteins fold into specific 3D structures, why lipid bilayers spontaneously assemble in water, and why complex organisms constantly radiate heat to offset the immense internal order they maintain.
Slide 9: Synthesizing Spontaneity in Bioenergetics
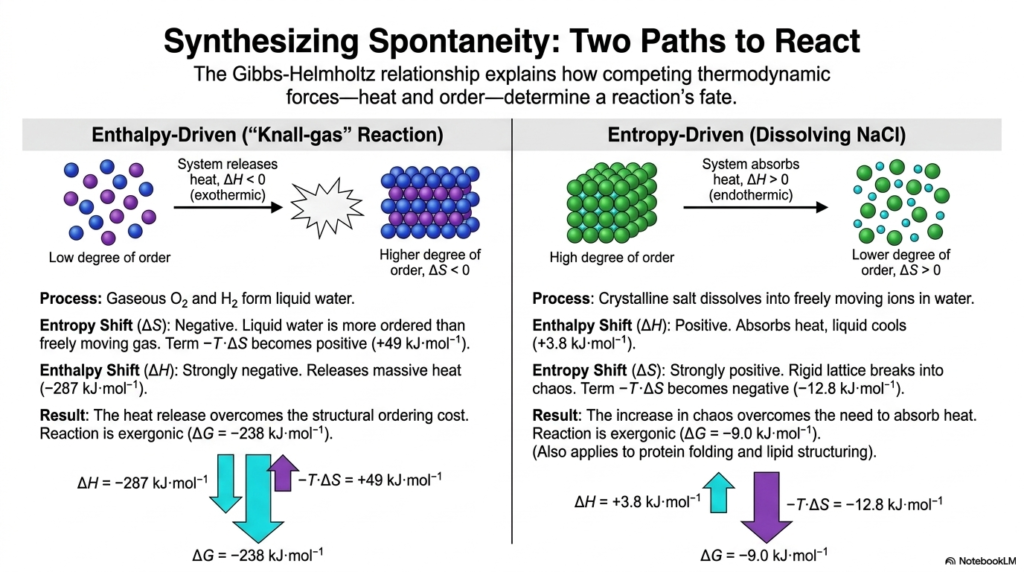
To truly grasp the Gibbs-Helmholtz relationship, we must look at how enthalpy and entropy compete in the real world to determine a reaction’s ultimate fate. This slide beautifully illustrates the two distinct paths to spontaneity studied in Bioenergetics: enthalpy-driven reactions and entropy-driven reactions.
First, consider the enthalpy-driven “Knall-gas” reaction, where gaseous oxygen and hydrogen combine to form liquid water. The formation of liquid water is inherently more ordered than a freely moving gas, resulting in an unfavorable negative shift in entropy. By the rules of Bioenergetics, this should stop the reaction dead. However, the reaction releases a massive amount of heat. This overwhelming heat release entirely overrides the structural ordering cost, resulting in a highly exergonic, spontaneous reaction.
On the other hand, look at the entropy-driven process of dissolving table salt (NaCl) in water. When crystalline salt dissolves into freely moving ions, the system actually absorbs heat from the environment, making the water slightly colder. Despite this unfavorable endothermic shift, the rigid salt lattice breaks into total chaos, resulting in a massively positive entropy shift.
This sudden explosion of molecular disorder overcomes the need to absorb heat, making the dissolving process spontaneous. For students of Bioenergetics, this is a profound revelation. These same competing thermodynamic forces govern everything from the folding of complex globular proteins to the delicate structuring of cellular lipid membranes. Spontaneity in biology is never just about heat; it is always a delicate tug-of-war between the release of energy and the relentless march toward chaos.
Slide 10: Reaction Kinetics and Bioenergetics
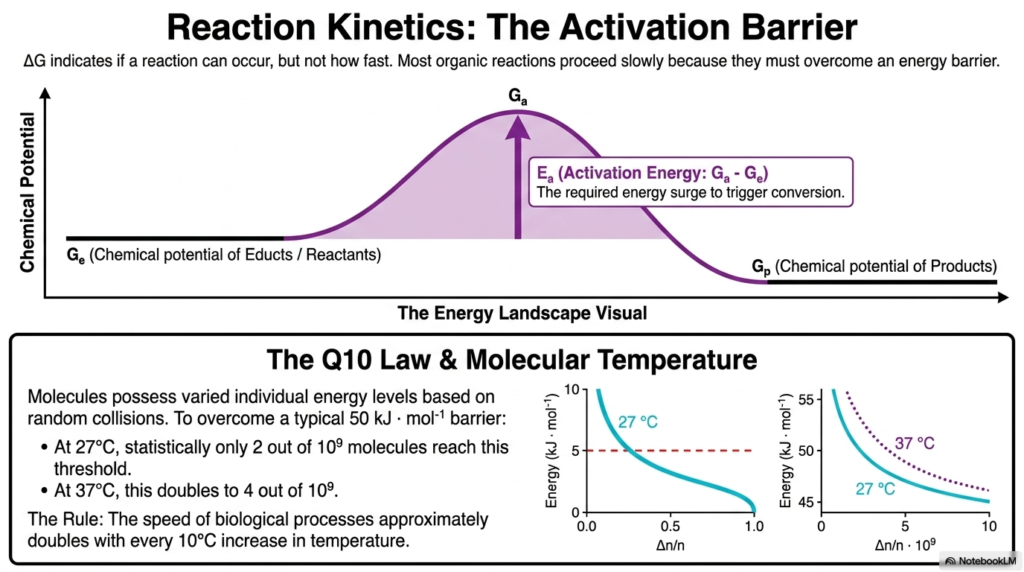
Up to this point, we have only discussed thermodynamics—whether a reaction can happen. But ΔG tells us absolutely nothing about how fast a reaction will happen. Welcome to the other half of Bioenergetics: Reaction Kinetics. Most organic reactions naturally proceed at an agonizingly slow pace because they are blocked by a massive energy barrier known as the Activation Energy.
As seen in the energy landscape visualization, molecules must absorb a sufficient energy threshold to trigger the conversion from reactants to products. Even if a reaction is highly spontaneous, it will not occur if the molecules cannot overcome this thermodynamic hill. Bioenergetics teaches us that molecules possess distinct energy levels determined strictly by random, chaotic collisions. According to the Q10 Law, at 27 degrees Celsius, only 2 out of every billion molecules would, on average, have enough kinetic energy to overcome the barrier. However, if you simply raise the molecular temperature by 10 degrees to 37 degrees Celsius, that number instantly doubles!
This beautifully straightforward rule demonstrates why the speed of biological processes approximately doubles with every 10-degree increase in temperature. Understanding the kinetics side of Bioenergetics is paramount for future clinicians. It explains why a severe fever can dangerously accelerate metabolic rates, why human bodies must maintain a strict core temperature of 37 degrees to keep enzymes functioning, and why refrigeration successfully slows bacterial growth. Thermodynamics may give the green light for a reaction to occur, but kinetics completely controls the biological speed limit.
Slide 11: Reaction Rates in Bioenergetics
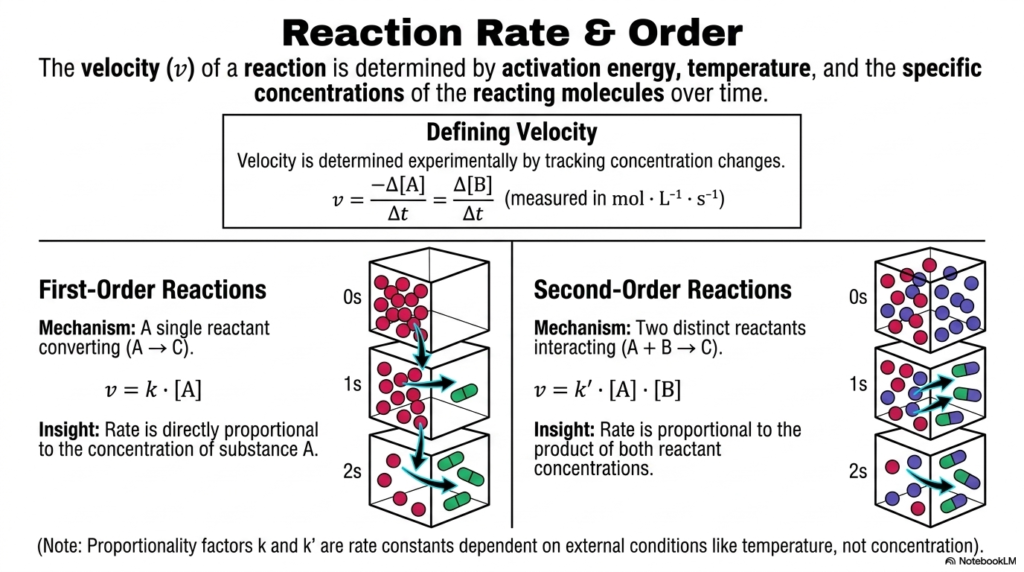
Diving deeper into kinetics, we must understand how to measure and predict the actual speed of molecular transformations. This slide focuses on reaction rates and reaction order, essential mathematical concepts in Bioenergetics. The overall velocity of any biochemical reaction is determined by a combination of the activation energy, the ambient temperature, and the concentrations of the reacting molecules over time.
Velocity is determined experimentally by tracking exactly how fast reactant concentrations drop or product concentrations rise. In Bioenergetics, we categorize these speeds into distinct reaction orders. A First-Order Reaction involves a single reactant spontaneously converting into a product. Here, the rate of the reaction is directly and linearly proportional to the concentration of that single substance. If you double the amount of the reactant, you exactly double the rate of the conversion.
Alternatively, a Second-Order Reaction requires that two distinct reactants physically collide and interact to form a product. The velocity here is far more complex, as it is proportional to the product of the concentrations of both reactants. These rates are linked by proportionality factors known as rate constants, which vary widely with external conditions such as temperature. For students studying Bioenergetics, calculating reaction orders is not just busy work; it is the fundamental basis of pharmacokinetics. When a doctor administers a drug, its clearance from the bloodstream follows these exact kinetic principles, ensuring you understand exactly how long a drug will remain active in a patient.
Slide 12: The Golden Rules of Bioenergetics

We have reached the grand synthesis. The final slide brilliantly summarizes the two distinct scientific domains that govern the transformation of matter in living systems. To master Bioenergetics, you must recognize that your cells are simultaneously navigating the rigid rules of Thermodynamics and the dynamic rules of Reaction Kinetics. They are two sides of the exact same biological coin.
Thermodynamics represents the “If” and the “How Much.” Its governing metric is the free enthalpy change (ΔG), which absolutely determines the spontaneity and equilibrium direction of any reaction. Influenced heavily by heat and order, it uses energetic coupling as its primary biological tool. As Bioenergetics demonstrates, the cell survives by using the large energy deficit of ATP hydrolysis to forcibly drive otherwise impossible, endergonic reactions forward. You cannot break the laws of thermodynamics, but you can cleverly couple them to your advantage.
Conversely, Reaction Kinetics represents the “When” and the “How Fast.” Its governing metrics are activation energy and velocity, determining the actual, real-world time scale of molecular transformation. While thermodynamics cannot be cheated, cells masterfully manipulate kinetics through Enzymatic Catalysis. By deploying specialized proteins to artificially lower the activation energy barrier, the body ensures that spontaneous reactions occur at biologically viable, lightning-fast speeds. Ultimately, Bioenergetics is the beautiful marriage of these two domains. Without thermodynamics, there is no energy to sustain life. Without kinetics and enzymes, life would happen too slowly to matter. By understanding both, you have cracked the ultimate code of cellular biology.
Check out our social media channels:
Please read our Content Disclaimer Statement.