6. Water and pH: Exploring the Biochemical Foundations of Life
You are mostly water. We hear this fact all the time, but have you ever stopped to think about what that actually means for the microscopic machinery inside your cells? You might be used to thinking of water as just a passive liquid that fills up a cup, or the quiet background fluid inside a cell where the “real” biology happens. The truth is far more exciting. Water is the primary actor on the biochemical stage.
The core purpose of this slide deck is to break down the profound driving forces of aqueous biochemistry. For college and medical school students, this is where rote memorization ends and true physiological understanding begins. We are going to explore how the physical properties of a simple solvent dictate protein folding, membrane formation, and enzyme-catalyzed reactions that make your heartbeat possible. Let’s dive into the biological matrix!
Slide 1: The Biochemistry of Aqueous Systems: A Masterclass in Water and pH

Welcome to the foundation of life. When we dive into the study of cellular life, we are ultimately exploring the profound impact of water and pH on biological systems. You might be used to thinking of water as merely a solvent, but as this introductory slide illustrates, water is the primary actor on the biochemical stage. It is impossible to separate a cell’s structural integrity from the dynamics of water and pH.
This title slide, featuring a complex geometric tetrahedron, represents the hidden structure of aqueous systems. We are stepping away from the simplified view of biology and diving deep into the molecular dynamics, thermodynamics, and the crucial acid-base equilibria that govern human physiology. For college and medical school students, this is where rote memorization ends and true physiological understanding begins. This slide deck sets the stage for our journey into how water and pH govern molecular dynamics, thermodynamics, and the delicate equilibria that sustain life.
In biochemistry, context is everything. The proteins, carbohydrates, lipids, and nucleic acids that we study do not exist in a vacuum; they exist in an aqueous environment that constantly pushes, pulls, and shapes them. By mastering the concepts introduced in this presentation, you will build a rock-solid foundation for everything from pharmacology to pathophysiology. You will understand why certain drugs can cross cell membranes while others cannot, and why a slight shift in your blood’s acidity can be a life-threatening medical emergency. Let’s unpack the secrets of the biological matrix!
Slide 2: The Dipolar Molecule That Drives Water and pH Dynamics
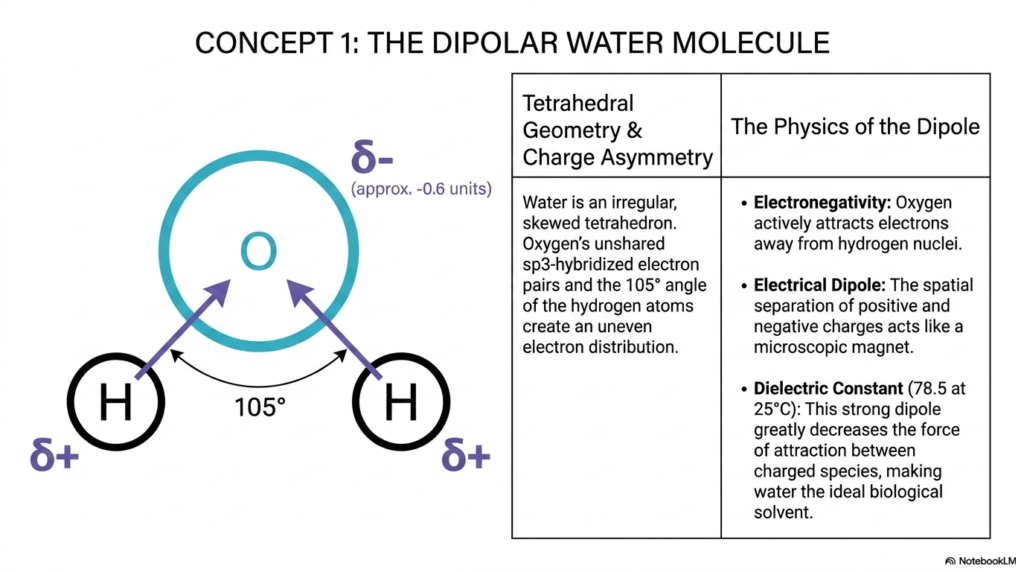
To truly grasp the complex behaviors of water and pH, we must first look at the unique geometry of the water molecule itself. On this slide, we see a detailed illustration of water represented as an irregular, skewed tetrahedron. The oxygen atom sits at the center, holding onto unshared sp3-hybridized electron pairs. Because the two hydrogen atoms are pushed together at a specific 105-degree angle, the electron distribution is heavily skewed. Oxygen is highly electronegative, meaning it strongly pulls the shared electrons away from the hydrogen nuclei.
This uneven sharing creates a charge asymmetry. The oxygen pole becomes partially negative, while the hydrogen poles become partially positive. This simple spatial arrangement is the very foundation of why water and pH play such a commanding role in biological solvent interactions. The spatial separation of these charges turns every single water molecule into a microscopic magnet, also known as an electrical dipole.
Because of this strong dipole, water possesses an exceptionally high dielectric constant of 78.5 at room temperature. In biochemistry, this means water is uniquely capable of neutralizing attractions between other charged molecules. This greatly decreases the force of attraction between external charged species, making it the ideal biological solvent. Understanding this dipole is your first major step in mastering the intricacies of water and pH in biochemistry. Without this fundamental geometric asymmetry, the solvent properties that allow ions to freely circulate in our bloodstream would not exist. Every metabolic pathway in your body, from muscle contraction to neuronal firing in your brain, relies entirely on this dipolar nature.
Slide 3: Hydrogen Bonding Networks: The Physical Backbone of Water and pH

As we continue exploring the physical chemistry of water and pH, we encounter the phenomenon of hydrogen bonding. The previous slide showed us that water is a dipole. Because opposite charges attract, the partially positive hydrogen atom of one water molecule is electrostatically drawn to the partially negative oxygen atom of a neighboring molecule. This creates a transient, flickering connection known as a hydrogen bond.
While each individual hydrogen bond is incredibly brief—having a half-life of merely a few picoseconds—they are constantly breaking and reforming. On average, each water molecule associates with approximately 3.5 others, creating a massive, interconnected tetrahedral lattice. The fact that a single molecule can bond with up to 3.5 others explains why the thermodynamic properties of water and its pH are so drastically different from those of molecules like methane.
Look at the comparison table provided on the slide. Water and methane have relatively similar molecular masses, but their physical states on Earth could not be more different. Because methane lacks a strong dipole moment, it cannot form hydrogen bonds. As a result, it boils at a freezing -47.6 degrees Celsius and exists as a gas. Water, on the other hand, requires a substantial input of heat energy to disrupt its extensive hydrogen-bond lattice, allowing it to remain liquid up to 100 degrees Celsius.
Ultimately, the stability of these hydrogen bond networks is what allows water and pH to maintain the liquid environment required for life on Earth. Understanding this thermodynamic resilience helps explain how the human body regulates temperature through sweating and why our internal environment remains incredibly stable.
Slide 4: Hydration and Solvation: How Water and pH Govern Solubility
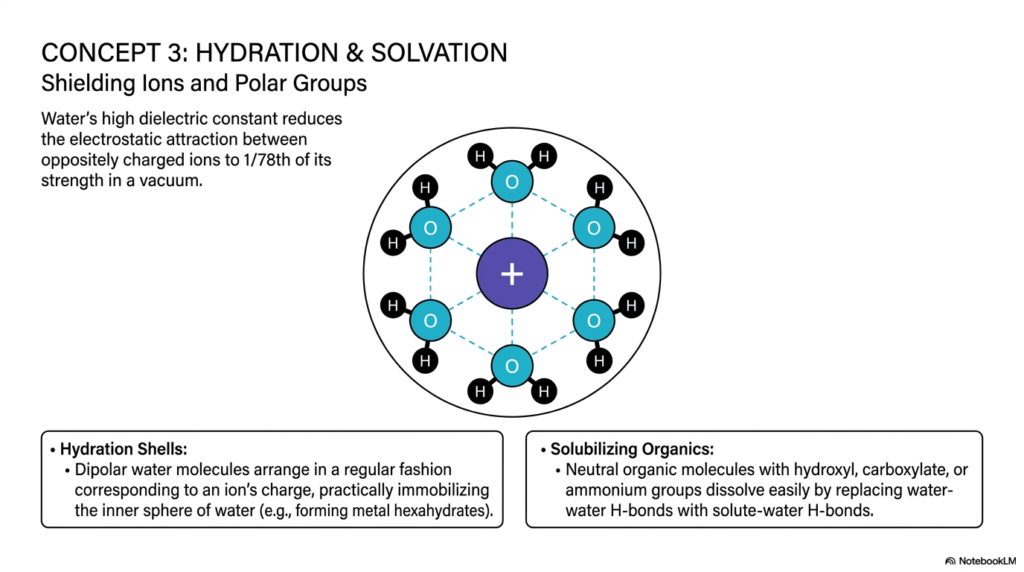
The interactions between water and pH become particularly relevant when we consider how substances dissolve in our cells. This slide tackles the concepts of hydration and solvation, revealing exactly how water shields ions and polar groups. Because water is a powerful dipole with a high dielectric constant, it reduces the electrostatic attraction between oppositely charged ions to just 1/78th of what their strength would be in a pure vacuum.
When a charged ion, such as a positively charged sodium ion, enters an aqueous environment, the dipolar water molecules immediately swarm it. They arrange themselves in a highly regular, ordered fashion corresponding to the ion’s charge. The partially negative oxygen atoms face inward toward the positive ion, forming a protective hydration shell. The high dielectric constant of water and its pH chemistry allow water to substantially reduce the electrostatic attraction between oppositely charged ions, effectively immobilizing the inner sphere of water and forming structures such as metal hexahydrates.
But water doesn’t just dissolve charged salts; it also solubilizes neutral organic molecules. Biologically crucial molecules containing hydroxyl, carboxylate, or ammonium groups dissolve easily because they can replace water-water hydrogen bonds with solute-water hydrogen bonds. When we study the dissolution of biological organics, it always circles back to the underlying rules of water and pH. If our biological solvent lacked this ability to form hydration shells, vital nutrients would precipitate out of our blood, and cellular transport would immediately cease. Recognizing how water cages and shields these molecules gives us critical insight into modern drug design and systemic delivery.
Slide 5: The Hydrophobic Effect: Entropy, Water and pH
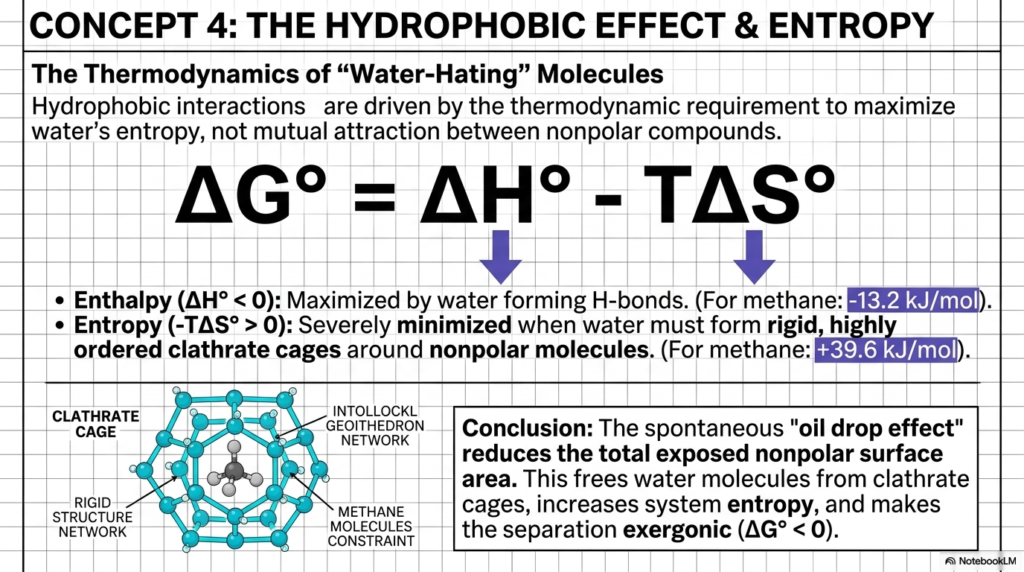
The thermodynamic principles of water and pH extend far beyond simple dissolving; they drive the complex folding of proteins through the hydrophobic effect. Have you ever wondered why oil and water refuse to mix, or how a protein knows exactly how to fold into its functional three-dimensional shape? The answer lies not in an active repulsion between water and nonpolar molecules, but in the universe’s relentless drive toward chaos, known in physics as entropy.
When a “water-hating” or hydrophobic molecule like methane enters an aqueous environment, water molecules cannot form hydrogen bonds with it. Instead, they are forced to form highly ordered, rigid structures called clathrate cages around the nonpolar intruder. This significantly reduces the water’s entropy, which is thermodynamically unfavorable. To fix this, nonpolar molecules spontaneously clump together. This spontaneous clumping, driven by the thermodynamic rules of water and pH, minimizes the exposed nonpolar surface area.
By bunching up, hydrophobic molecules free the surrounding water molecules from their rigid clathrate cages, thereby dramatically increasing the system’s entropy. The slide illustrates this beautifully with the Gibbs Free Energy equation, showing that this separation is an exergonic, spontaneous process. By studying this “oil drop effect,” we see that the laws of water and pH dictate the architecture of life through the pursuit of maximum entropy. This thermodynamic mechanism forces the hydrophobic side chains of amino acids into the interior of a folding protein, protecting them from the aqueous environment. Without this entropy-driven phenomenon, the functional enzymes necessary for human survival would simply unravel.
Slide 6: Amphipathic Assemblies: Membrane Formation in Water and pH
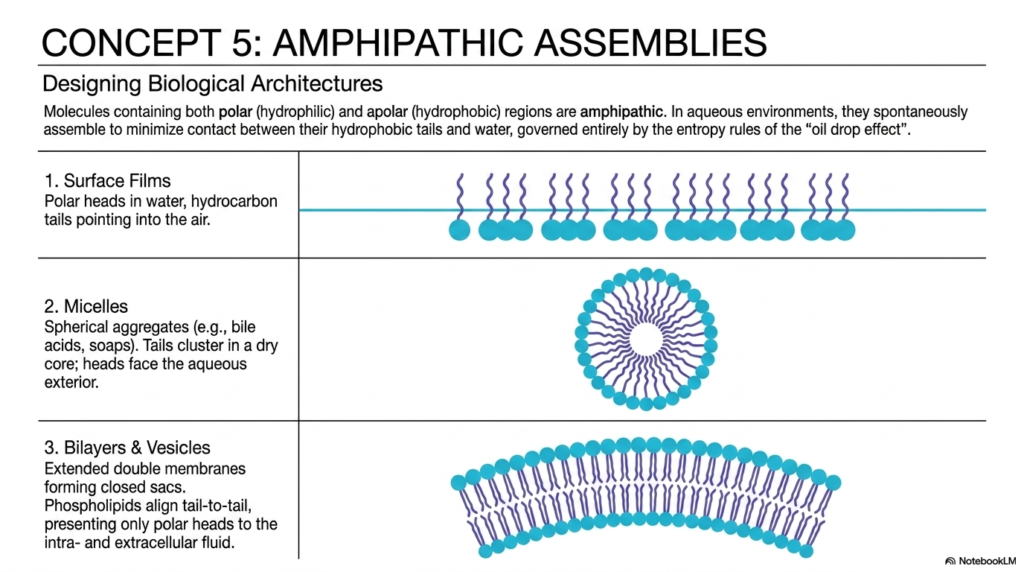
The unique chemistry of water and pH is perfectly illustrated by amphipathic molecules, which possess both water-loving and water-fearing regions. Amphipathic molecules are the ultimate biological multitaskers. Because they have a hydrophilic polar head and a hydrophobic nonpolar tail, they face a thermodynamic dilemma when placed in an aqueous environment. They must find a way to satisfy both regions simultaneously without violating the rules of entropy.
In the context of water and pH, these molecules spontaneously organize into structures such as surface films, micelles, and lipid bilayers to minimize unfavorable thermodynamic interactions. The slide breaks down three primary architectures. First, they can form surface films at the air-water interface, where polar heads touch the water, and nonpolar tails face the air. Second, they can form micelles, which are spherical structures in which the nonpolar tails are hidden inside a core, completely sequestered from the surrounding liquid.
Finally, and most importantly for college and medical students, they form bilayers and vesicles. In a bilayer, two sheets of amphipathic molecules align so their nonpolar tails face each other, forming a central hydrophobic core. Without the driving forces dictated by water and pH, the lipid bilayer membranes that enclose every single cell in your body simply could not exist. These membranes create vital biological compartmentalization, allowing cells to maintain distinct internal environments. They act as selective barriers, ensuring that the precise chemical concentrations required for cellular respiration and action potentials are rigorously protected from the chaotic external environment.
Slide 7: Noncovalent Forces: The Hidden Stabilizers of Water and pH

While strong covalent bonds dictate the primary sequence of molecules, the rules of water and pH govern the noncovalent forces that dictate 3D architecture. Think of covalent bonds as the tough, unbreakable spine of a biological molecule—like the sequence of amino acids in a polypeptide chain. However, a straight chain is biologically useless. To become a functional tool, like an enzyme or a cellular receptor, the molecule must fold, twist, and lock into a highly specific three-dimensional shape.
This slide beautifully details the four major types of noncovalent interactions. We have Hydrogen Bonds, which share unshielded protons and are highly directional. We have Salt Bridges, which are electrostatic interactions between oppositely charged groups that facilitate binding over larger distances. We also see the Hydrophobic Effect, which is a systematic thermodynamic effect driven by maximizing entropy. Finally, there are van der Waals forces—extremely weak attractions between transient dipoles that become incredibly powerful when multiplied across a massive macromolecule.
Understanding the water environment and pH is crucial because forces such as hydrogen bonds, salt bridges, and van der Waals interactions depend heavily on the surrounding solvent. Water directly competes with these forces, attempting to form its own hydrogen bonds with the biomolecules. Ultimately, the functional competence of DNA, proteins, and enzymes is entirely dependent on the delicate balance of noncovalent forces in water and at pH. The DNA double helix is the perfect example; the two strands of your genetic code are held together not by permanent bonds, but by delicate hydrogen bonds.
Slide 8: Water as a Nucleophile: Reactivity in Water and pH
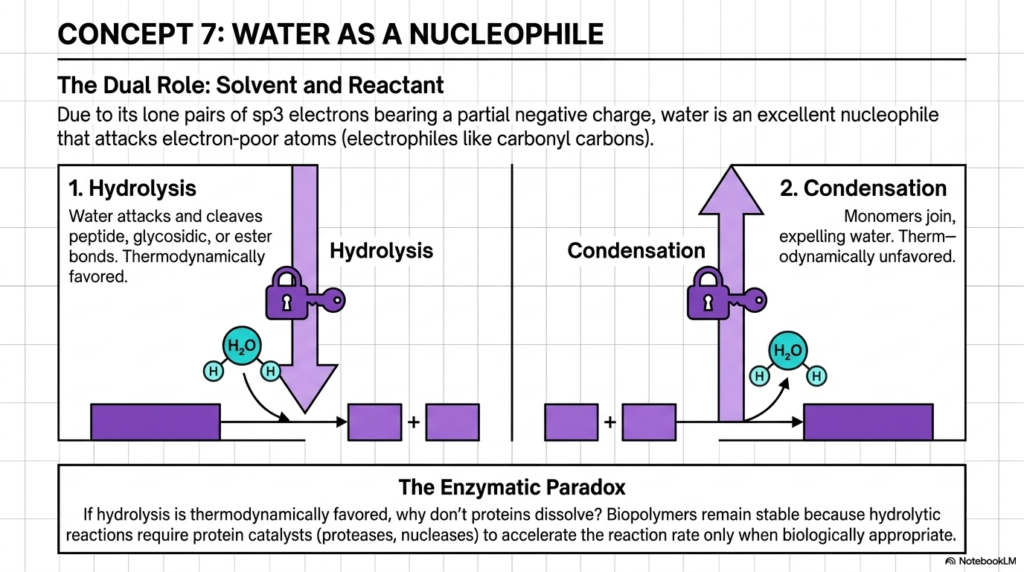
It is crucial to remember that in the study of water and pH, water is not just a passive background solvent; it is an active, highly potent reactant. So far, we’ve discussed how water shapes molecules and provides a medium for them to float in. However, this slide shifts our focus strictly to water’s chemical reactivity. Due to its lone pairs of sp3 hybridized electrons that carry a partial negative charge, water acts as an excellent nucleophile—meaning it actively seeks out and attacks electron-poor atoms.
This nucleophilic nature leads to two highly significant biological reactions. First is hydrolysis, where water attacks and cleaves peptide, glycosidic, or ester bonds. This reaction is thermodynamically favored, meaning nature inherently wants to break these molecules apart. Conversely, we have condensation, in which monomers join to expel a water molecule, a thermodynamically unfavorable process. The lone pairs of electrons on the oxygen atom make water an excellent nucleophile, directly linking water and pH to enzymatic cleavage and hydrolysis.
This brings us to a fascinating question raised at the bottom of the slide: The Enzymatic Paradox. If hydrolysis is thermodynamically favored, why don’t the proteins in our body just spontaneously dissolve into mush? The enzymatic paradox highlights why the regulatory environment of water and pH is so essential for preventing our biopolymers from spontaneously dissolving. While water inherently wants to break these bonds, the reaction happens far too slowly on its own. Our biopolymers remain stable because hydrolytic reactions require specific protein catalysts to accelerate the reaction only when biologically appropriate.
Slide 9: The Ionization of Water: The Core of Water and pH

We finally reach the central mathematical concept linking the dual nature of water and pH: the slight tendency of water to ionize. While the vast majority of water molecules remain intact, a very tiny fraction naturally dissociates into a positively charged hydronium ion and a negatively charged hydroxide ion. This happens through rapid, intermolecular proton transfer.
This intermolecular proton transfer is the fundamental physical process that makes the study of water and pH possible, thereby establishing a constant ionic product. The slide breaks down the math perfectly. At any given instant, the probability that a hydrogen atom exists as a free ion is incredibly small—just 1.8 x 10^-9. However, because pure water is incredibly concentrated at 55.56 moles per liter, we multiply the probability by the concentration. This reveals that the concentration of protons in pure water is exactly 1.0 x 10^-7 mol/L.
Because the concentration of water itself is so vast that it remains effectively constant, biochemists incorporate it into a standard dissociation constant, creating the Ion Product of Water. The equation Kw = 1.0 x 10^-14 is the fundamental mathematical baseline for all acid-base chemistry in the human body. Mastering this protonic probability is absolutely non-negotiable for medical and college students trying to conquer the chemistry of water and pH. This constant dictates that if the proton concentration in a cellular fluid increases, the hydroxide concentration must decrease proportionally to maintain the same equilibrium, which is vital for clinical lab interpretations.
Slide 10: Biological Compartments: The Scale of Water and pH

The logarithmic measurement created by Sörensen bridges the gap between pure chemistry and the physiological realities of water and pH. Working with tiny numbers in scientific notation is mathematically tedious. To simplify this, the pH scale was introduced as the negative logarithm of the hydrogen ion concentration. This means a difference of just 1 on the pH scale represents a massive 10-fold change in actual proton concentration.
By examining different biological compartments, we see that the regulation of water and pH is not uniform throughout the body. The slide provides a phenomenal visual map of our internal compartments. Starting at the highly acidic end, we have gastric juice with a pH around 2.0, providing the extreme acidity necessary to break down food. Moving up the scale, we find lysosomes, the cell’s recycling centers, operating at a pH of 4.5 to 5.5. This ensures that if a lysosome ruptures, its destructive enzymes will harmlessly deactivate in the neutral cytoplasm.
Whether it is the highly acidic environment of a lysosome or the strictly regulated blood plasma, the local balance of water and pH dictates specific organelle functions. Our cellular cytoplasm sits at a neutral baseline of 7.0-7.3. However, our blood plasma operates in a critically narrow margin of 7.35 to 7.45. Even a slight deviation from this tight window results in life-threatening medical conditions known as acidosis or alkalosis. This compartmentalization demonstrates that our bodies actively engineer specific chemical environments to drive distinct metabolic tasks.
Slide 11: Weak Acids, Bases, and pKa in Water and pH

In biological systems, the interactions of water and pH are largely governed by weak acids and bases that only partially dissociate. Unlike strong acids, which completely shatter into ions the moment they touch liquid, essential biochemicals act much more conservatively. Key biological functional groups—such as carboxyls, amines, and phosphates—exist in equilibrium, constantly shuttling between their protonated and deprotonated conjugate base states.
The concept of pKa is intricately tied to water and pH, describing the exact dissociation constant and acid strength of essential biochemicals. Just as pH uses a logarithmic scale to describe proton concentration, pKa uses a negative logarithm to describe an acid’s dissociation constant. The rule is simple: the lower the pKa, the stronger the acid and the more eagerly it wants to give up its proton.
We must also recognize that structural influences, such as adjacent charges, alter how these molecules behave in the context of water and pH. This slide highlights two key environmental factors that influence an acid’s strength. First, an adjacent negative charge will hinder proton release due to electrostatic repulsion. Second, the dielectric medium plays a massive role. If a weak acid is moved from the aqueous exterior of a protein into the nonpolar, hydrophobic interior, its pKa shifts radically because the nonpolar environment cannot efficiently solvate charged species. For pharmacology students, this concept dictates how effectively a drug will be absorbed across different tissues.
Slide 12: The Henderson-Hasselbalch Equation: Calculating Water and pH
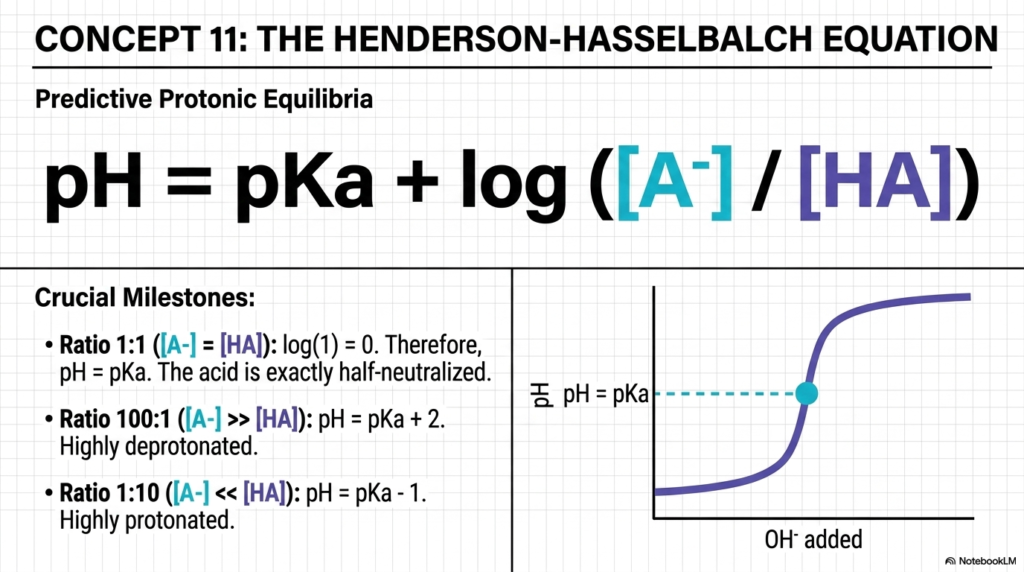
The Henderson-Hasselbalch equation is the ultimate predictive tool for mastering the mathematical relationship between water and pH. It is nearly impossible to pass a college biochemistry or medical physiology exam without understanding this formula. This equation brilliantly connects environmental acidity to a molecule’s intrinsic acid strength and the ratio of its unprotonated to protonated forms.
By charting the ratio of a conjugate base to its weak acid, students can pinpoint exactly how water and pH will shift during titrations. The slide outlines three crucial mathematical milestones you must memorize. First, when the concentration of the conjugate base perfectly equals the weak acid, the ratio is 1:1. The log of 1 is exactly 0. Therefore, at this exact moment, pH equals pKa, meaning the acid is exactly half-neutralized. Second, if the ratio is 100:1, the molecule is highly deprotonated. Third, if the ratio is flipped to 1:10, the molecule is highly protonated.
Understanding these crucial milestones, such as when the ratio is exactly 1:1, demystifies the complex behavior of buffers in water and pH biochemistry. The sigmoidal curve illustrated on the slide is a classic titration curve. The flat middle section of the curve represents the zone where adding base causes the least change in overall pH. This visual shows that molecules are highly resistant to environmental changes when they hover near their specific pKa values, enabling clinicians to precisely correct dangerous chemical imbalances.
Slide 13: Physiological Buffer Systems: Defending Water and pH
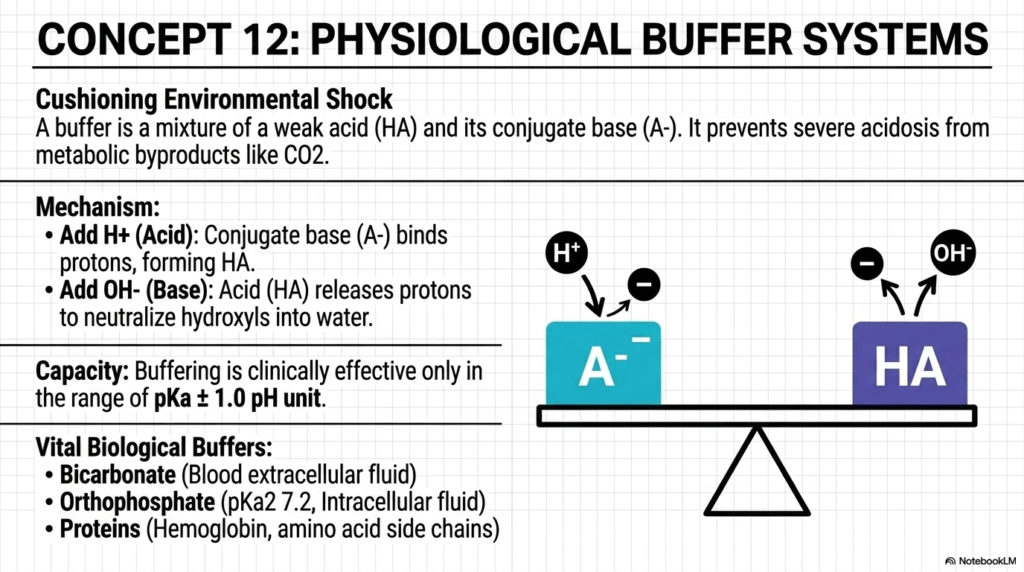
To cope with the constant production of metabolic byproducts, our bodies rely on buffer systems to absorb the environmental shocks caused by water and pH changes. Every single time you flex a muscle, digest a meal, or take a breath, your cells generate acidic byproducts like carbon dioxide. If left unchecked, these protons would flood your bloodstream and denature your proteins. A buffer—a carefully balanced mixture of a weak acid and its conjugate base—acts as a chemical shock absorber.
The mechanism is beautifully elegant. If a sudden influx of acid threatens the system, the conjugate base aggressively binds the free protons, converting them back into the weak acid. Conversely, if a sudden influx of base appears, the weak acid willingly sacrifices its proton to neutralize the threat into harmless water. The clinical effectiveness of these buffers depends entirely on the principles of water and pH, specifically operating within one unit of their pKa. This means a buffer is only functionally useful when the environmental acidity is within a tight range of pKa ± 1.0.
Vital systems, such as the bicarbonate buffer in our blood, demonstrate the life-saving physiological role of water and pH balance. The slide outlines the major biological defenders. The bicarbonate system works tirelessly in our extracellular fluid to neutralize metabolic acids. Inside the cell, the orthophosphate system takes over to protect the intracellular fluid. Additionally, the countless proteins in our body act as massive, complex buffering agents to ensure survival.
Slide 14: The Aqueous Biochemical Loop: Synthesizing Water and pH
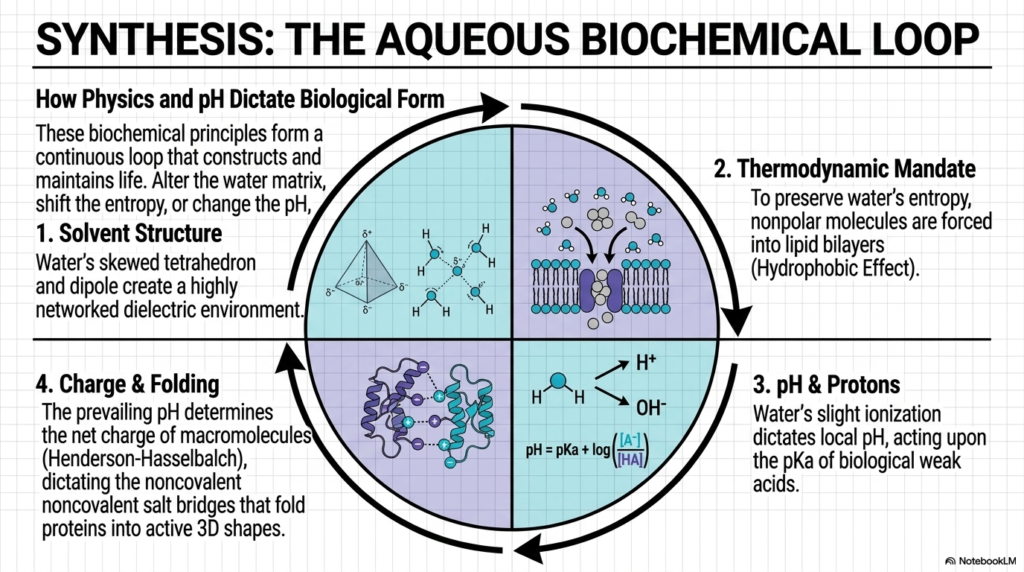
This final slide brings everything full circle, demonstrating how the fundamental physics of water and pH dictate all biological form and function. Biochemistry is not a series of disconnected facts; it is an integrated, continuous loop of cause and effect. This synthesis diagram visually unites the four major pillars we have explored throughout this presentation: solvent structure, thermodynamic mandates, protonic equilibria, and the resulting charge and folding of macromolecules.
From the thermodynamic mandate of protein folding to the precise solvent structures of the lipid bilayer, the rules of water and pH are inescapable. The physical shape of the water molecule and its hydrogen-bonding network establish the foundational solvent structure. This physical reality forces nonpolar molecules to hide, creating the thermodynamic mandate that drives the formation of protective lipid bilayers and dictates the intricate folding of enzymes. Simultaneously, water’s slight tendency to ionize creates the baseline for protonic behavior, introducing the Henderson-Hasselbalch equation.
As you move forward in your biochemical education, remember that this continuous loop of water and pH interactions is the core engine of cellular life. You cannot change one quadrant of this loop without instantly affecting the others. A shift in thermodynamics alters folding; a shift in acidity alters molecular charge. By grasping this holistic synthesis, college and medical school students transition from merely memorizing metabolic pathways to truly understanding the magnificent, dynamic, and beautifully complex chemical logic that makes human life possible.
Check out our social media channels:
Please read our Content Disclaimer Statement.